Лекция
Привет, Вы узнаете о том , что такое Уравнения Лотки–Вольтерры, Разберем основные их виды и особенности использования. Еще будет много подробных примеров и описаний. Для того чтобы лучше понимать что такое Уравнения Лотки–Вольтерры , настоятельно рекомендую прочитать все из категории Моделирование и Моделирование систем.
Уравнения Лотки–Вольтерры , также известные как модель хищник–жертва Лотки–Вольтерры , представляют собой пару нелинейных дифференциальных уравнений первого порядка , часто используемых для описания динамики биологических систем, в которых взаимодействуют два вида: один как хищник , а другой как жертва. Численность популяций изменяется со временем в соответствии с этой парой уравнений: ,
где
 и
и  представляют собой мгновенные темпы роста двух популяций;
представляют собой мгновенные темпы роста двух популяций;Решение дифференциальных уравнений является детерминированным и непрерывным . Это, в свою очередь, означает, что поколения хищника и жертвы постоянно перекрываются.
Система уравнений Лотки-Вольтерры является примером модели популяции Колмогорова (не путать с более известными уравнениями Колмогорова ) , которая представляет собой более общую структуру, позволяющую моделировать динамику экологических систем с взаимодействиями хищник-жертва, конкуренцией , болезнями и мутуализмом .
Предполагается, что жертвы имеют неограниченный запас пищи и размножаются экспоненциально, если не подвергаются хищничеству; этот экспоненциальный рост представлен в приведенном выше уравнении членом αx . Предполагается, что скорость хищничества по отношению к жертвам пропорциональна скорости встречи хищников и жертв; это представлено выше членом βxy . Если x или y равны нулю, то хищничества не может быть. С помощью этих двух членов приведенное выше уравнение для жертв можно интерпретировать следующим образом: скорость изменения популяции жертв определяется ее собственной скоростью роста минус скорость, с которой на нее охотятся.
Термин δxy обозначает рост популяции хищников. (Обратите внимание на сходство со скоростью хищничества; однако используется другая константа, поскольку скорость роста популяции хищников не обязательно равна скорости потребления ими добычи). Термин γy обозначает скорость сокращения численности хищников из-за естественной смерти или эмиграции; в отсутствие добычи это приводит к экспоненциальному спаду. Таким образом, уравнение показывает, что скорость изменения популяции хищников зависит от скорости потребления ими добычи за вычетом их собственной скорости смертности.
Модель хищник-жертва Лотки-Вольтерры предполагает ряд допущений относительно окружающей среды и биологии популяций хищника и жертвы:
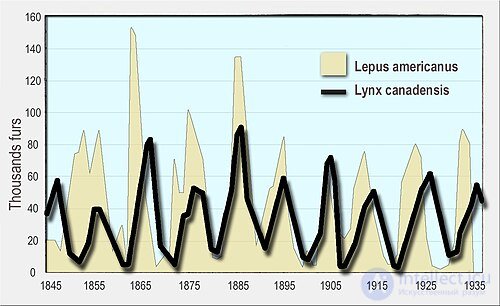
Количество меха зайца-беляка (желтый, на заднем плане) и канадской рыси (черная линия, на переднем плане), проданного компании «Гудзонов залив» . Канадские рыси питаются зайцами-беляками.
Ни одно из вышеперечисленных предположений, вероятно, не будет выполняться для естественных популяций. Тем не менее, модель Лотки-Вольтерры демонстрирует два важных свойства популяций хищников и жертв, и эти свойства часто распространяются на варианты модели, в которых эти предположения ослаблены:
Во-первых, динамика популяций хищников и жертв имеет тенденцию к колебаниям. Колебания численности хищников и жертв наблюдались в естественных популяциях, например, данные по рыси и зайцу-беляку компании Hudson's Bay Company и популяции лосей и волков в национальном парке острова Ройал .
Во-вторых, равновесие популяции в этой модели обладает свойством, что равновесная плотность добычи (задается формулой)  ) зависит от параметров хищника и равновесной плотности хищника (определяемой формулой
) зависит от параметров хищника и равновесной плотности хищника (определяемой формулой  ) на параметры добычи. Это приводит, как следствие, к увеличению, например, темпов роста добычи.α Это приводит к увеличению равновесной плотности хищников, но не равновесной плотности жертв. Улучшение условий для жертв приносит пользу хищнику, а не жертве (это связано с парадоксом пестицидов и парадоксом обогащения ). Демонстрацией этого явления служит увеличение процента пойманной хищной рыбы в годы Первой мировой войны (1914–1918), когда темпы роста жертв возросли из-за сокращения рыболовных усилий.
) на параметры добычи. Это приводит, как следствие, к увеличению, например, темпов роста добычи.α Это приводит к увеличению равновесной плотности хищников, но не равновесной плотности жертв. Улучшение условий для жертв приносит пользу хищнику, а не жертве (это связано с парадоксом пестицидов и парадоксом обогащения ). Демонстрацией этого явления служит увеличение процента пойманной хищной рыбы в годы Первой мировой войны (1914–1918), когда темпы роста жертв возросли из-за сокращения рыболовных усилий.
Еще один пример — экспериментальное удобрение океана железом . В нескольких экспериментах в океане растворяли большие количества солей железа. Ожидалось, что железо, являющееся лимитирующим питательным веществом для фитопланктона, будет стимулировать рост фитопланктона и поглощать углекислый газ из атмосферы. Добавление железа обычно приводит к кратковременному цветению фитопланктона, который быстро потребляется другими организмами (такими как мелкая рыба или зоопланктон ) и ограничивает эффект обогащения главным образом увеличением плотности хищников, что, в свою очередь, ограничивает поглощение углерода . Это соответствует предсказаниям равновесных плотностей популяций модели хищник-жертва Лотки-Вольтерры и является особенностью, которая сохраняется и в более сложных моделях, в которых ослабляются ограничительные предположения простой модели.
Модель Лотки-Вольтерры имеет дополнительные применения в таких областях, как экономика и маркетинг . Она может быть использована для описания динамики рынка с несколькими конкурентами, взаимодополняющими платформами и продуктами, экономикой совместного потребления и многим другим. Существуют ситуации, когда один из конкурентов вытесняет других конкурентов с рынка, и другие ситуации, когда рынок достигает равновесия, в котором каждая фирма стабилизирует свою долю рынка. Об этом говорит сайт https://intellect.icu . Также возможно описание ситуаций, когда в отрасли происходят циклические изменения или хаотические ситуации без равновесия, а изменения частые и непредсказуемые.
В экономике кривая Филлипса , показывающая статистическую взаимосвязь между безработицей и темпом изменения номинальной заработной платы, была связана с моделью Гудвина . Эта модель переосмысливает динамику биологического взаимодействия хищник-жертва, описанную моделью Лотки-Вольтерры, в экономических терминах. Способ взаимодействия двух видов в этой модели привел Гудвина к проведению параллелей с марксистским классовым конфликтом . Обобщение модели хищник-жертва Колмогорова, наряду с дальнейшим развитием модели Гудвина, расширило эти идеи.
Модель хищник-жертва Лотки-Вольтерры была впервые предложена Альфредом Дж. Лоткой в теории автокаталитических химических реакций в 1910 году. По сути, это было логистическое уравнение , первоначально выведенное Пьером Франсуа Верхульстом . В 1920 году Лотка расширил модель, через Андрея Колмогорова , на «органические системы», используя в качестве примера вид растений и вид травоядных животных , а в 1925 году он использовал уравнения для анализа взаимодействий хищник-жертва в своей книге по биоматематике . Тот же набор уравнений был опубликован в 1926 году Вито Вольтеррой , математиком и физиком, который заинтересовался математической биологией . Исследование Вольтерры было вдохновлено его общением с морским биологом Умберто Д'Анконой , который в то время ухаживал за его дочерью, а позже стал его зятем. Д'Анкона изучал улов рыбы в Адриатическом море и заметил, что процент пойманной хищной рыбы увеличился в годы Первой мировой войны (1914–1918). Это его озадачило, поскольку рыболовные усилия были значительно сокращены в годы войны, и, поскольку хищная рыба была предпочтительным уловом, интуитивно можно было ожидать увеличения процента хищной рыбы. Вольтерра разработал свою модель для объяснения наблюдения Д'Анконы и сделал это независимо от Альфреда Лотки. Он указал на более раннюю работу Лотки в своей публикации, после чего модель стала известна как «модель Лотки-Вольтерры».
Позже модель была расширена, чтобы включить зависимый от плотности рост добычи и функциональную реакцию в форме, разработанной К. С. Холлингом ; модель, которая стала известна как модель Розенцвейга-МакАртура. Как модель Лотки-Вольтерры, так и модель Розенцвейга-МакАртура использовались для объяснения динамики естественных популяций хищников и добычи.
В конце 1980-х годов появилась альтернатива модели хищник-жертва Лотки-Вольтерры (и ее обобщениям, зависящим от общей жертвы) — модель, зависящая от соотношения, или модель Ардити-Гинзбурга . [ 23 ] Достоверность моделей, зависящих от жертвы или соотношения, широко обсуждалась.
Уравнения Лотки-Вольтерры имеют долгую историю применения в экономической теории ; их первое применение обычно приписывают Ричарду Гудвину в 1965 или 1967 году .
Уравнения имеют периодические решения. Эти решения не имеют простого выражения в терминах обычных тригонометрических функций , хотя они вполне поддаются вычислению.
Если ни один из неотрицательных параметров α , β , γ , δ не равен нулю, три из них можно включить в нормализацию переменных, оставив только один параметр: поскольку первое уравнение однородно по x , а второе по y , параметры β / α и δ / γ могут быть включены в нормализации y и x соответственно, а γ — в нормализацию t , так что только α / γ остается произвольным. Это единственный параметр, влияющий на характер решений.
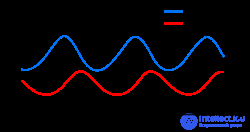
Динамика численности жертв и хищников с течением времени
Линеаризация уравнений дает решение, аналогичное простому гармоническому движению , где популяция хищников отстает от популяции жертв на 90° в цикле.
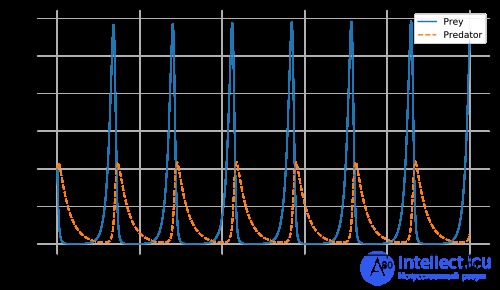
Отложим в сторону проблему динамики популяций кроликов и лис.
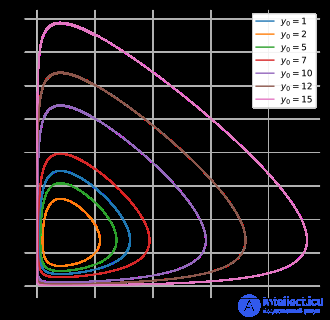
Диаграмма фазового пространства для задачи «хищник-жертва» при различных начальных условиях популяции хищников.
Предположим, есть два вида животных: кролик (жертва) и лиса (хищник). Если начальная плотность составляет 10 кроликов и 10 лис на квадратный километр, можно построить график изменения численности этих двух видов во времени, учитывая, что темпы роста и смертности кроликов составляют 1,1 и 0,4, а лис — 0,1 и 0,4 соответственно. Выбор временного интервала произволен.
Также можно параметрически отображать решения в виде орбит в фазовом пространстве , не представляя время, но с одной осью, представляющей количество добычи, и другой осью, представляющей плотность хищников для всех моментов времени.
Это соответствует исключению времени из двух приведенных выше дифференциальных уравнений, в результате чего получается одно дифференциальное уравнение.

Уравнение связывает переменные x (жертва) и y (хищник). Решения этого уравнения представляют собой замкнутые кривые. Оно поддается методу разделения переменных : интегрирование

приводит к неявному взаимоотношению
В=δх−γлн(х)+βй−αлн(й),
где V — постоянная величина, зависящая от начальных условий и сохраняющаяся на каждой кривой.
Примечание: Эти графики иллюстрируют серьезное потенциальное ограничение в применении в качестве биологической модели: при таком конкретном выборе параметров в каждом цикле популяция кроликов сокращается до крайне малых значений, но затем восстанавливается (в то время как популяция лис остается значительной при самой низкой плотности кроликов). Однако в реальных условиях случайные колебания дискретного числа особей могут привести к фактическому вымиранию кроликов, а следовательно, и лис. Эта проблема моделирования получила название «проблема атто-лисы», где атто- лиса — это условное 10⁻¹⁸ лис . Плотность 10⁻¹⁸ лис на квадратный километр соответствует в среднем примерно 5×10⁻¹⁰ лис на поверхности Земли, что на практике означает вымирание лис.
Поскольку количество  сохраняется во времени, играя роль гамильтоновой функции системы. Чтобы убедиться в этом, мы можем определить скобку Пуассона следующим образом
сохраняется во времени, играя роль гамильтоновой функции системы. Чтобы убедиться в этом, мы можем определить скобку Пуассона следующим образом  Тогда уравнения Гамильтона принимают следующий вид :
Тогда уравнения Гамильтона принимают следующий вид :  Переменныех
Переменныех ий
ий не являются каноническими, поскольку
не являются каноническими, поскольку  Однако, используя преобразования
Однако, используя преобразования  и
и  Мы пришли к канонической форме уравнений Гамильтона, включающей гамильтониан.
Мы пришли к канонической форме уравнений Гамильтона, включающей гамильтониан.  :
:  Скоба Пуассона для канонических переменных(q,п)
Скоба Пуассона для канонических переменных(q,п) теперь принимает стандартную форму
теперь принимает стандартную форму  .
.

Другой пример охватывает:
α = 2/3 , β = 4/3 , γ = 1 = δ . Предположим, что x и y измеряются тысячами. Кружки представляют начальные условия для жертвы и хищника от x = y = 0,9 до 1,8 с шагом 0,1. Фиксированная точка находится в (1, 1/2).
В модельной системе хищники процветают, когда добычи много, но в конечном итоге ее запасы превышают возможности ее пропитания, и ее численность сокращается. По мере снижения численности хищников популяция добычи снова увеличивается. Эта динамика продолжается в цикле роста и сокращения численности популяции .
В модели равновесие популяции достигается тогда, когда ни один из уровней популяции не изменяется, то есть когда обе производные равны 

Приведенная выше система уравнений дает два решения:  и
и 
Следовательно, существует два состояния равновесия.
Первое решение фактически представляет собой вымирание обоих видов. Если численность обеих популяций равна нулю, то она будет оставаться на этом уровне бесконечно. Второе решение представляет собой фиксированную точку, в которой обе популяции поддерживают свою текущую, ненулевую численность и, в упрощенной модели, делают это бесконечно. Уровни численности популяций, при которых достигается это равновесие, зависят от выбранных значений параметров α , β , γ и δ .
Устойчивость неподвижной точки в начале координат можно определить, выполнив линеаризацию с использованием частных производных .
Матрица Якоби модели «хищник-жертва» имеет вид:  и известна как матрица сообщества .
и известна как матрица сообщества .
При вычислении в стационарном состоянии (0, 0) матрица Якоби J принимает видДж.( 
Собственные значения этой матрицы равны 
В данной модели α и γ всегда больше нуля, и, следовательно, знак собственных значений всегда будет различаться. Таким образом, неподвижная точка в начале координат является седловой точкой .
Нестабильность этой неподвижной точки имеет важное значение. Если бы она была стабильной, к ней могли бы притягиваться ненулевые популяции, и, следовательно, динамика системы могла бы привести к вымиранию обоих видов во многих случаях начальных уровней популяций. Однако, поскольку неподвижная точка в начале координат является седловой точкой и, следовательно, нестабильной, отсюда следует, что вымирание обоих видов в данной модели затруднительно. (Фактически, это могло бы произойти только в том случае, если бы добыча была искусственно полностью истреблена, что привело бы к гибели хищников от голода. Если бы хищники были истреблены, популяция добычи росла бы неограниченно в этой простой модели.) Популяции добычи и хищника могут приблизиться к нулю на бесконечно малую величину и все же восстановиться.
Оценка J во второй неподвижной точке приводит 
Собственные значения этой матрицы равны 
Поскольку собственные значения являются чисто мнимыми и сопряжены друг другу, эта неподвижная точка должна быть либо центром замкнутых орбит в локальной окрестности, либо притягивающей или отталкивающей спиралью. В консервативных системах должны существовать замкнутые орбиты в локальной окрестности неподвижных точек, которые находятся в минимумах и максимумах сохраняемой величины. Сохраняемая величина, как показано выше, имеет вид:  на орбитах. Таким образом, орбиты вокруг неподвижной точки замкнуты и эллиптичны , поэтому решения периодические, колеблющиеся на малом эллипсе вокруг неподвижной точки с частотой.
на орбитах. Таким образом, орбиты вокруг неподвижной точки замкнуты и эллиптичны , поэтому решения периодические, колеблющиеся на малом эллипсе вокруг неподвижной точки с частотой.  и период
и период  .
.
Как показано на рисунке выше, циркулирующие колебания представляют собой замкнутые орбиты вокруг фиксированной точки: уровни популяций хищника и жертвы циклически меняются и колеблются без затухания вокруг фиксированной точки с определенной частотой.ω=αγ .
.
Значение постоянной движения V , или, эквивалентно,  , можно найти для замкнутых орбит вблизи неподвижной точки.
, можно найти для замкнутых орбит вблизи неподвижной точки.
Увеличение K приближает замкнутую орбиту к фиксированной точке. Наибольшее значение константы K получается путем решения задачи оптимизации. 
Таким образом, максимальное значение K достигается в стационарной (фиксированной) точке.  и составляет
и составляет  где e — число Эйлера .
где e — число Эйлера .
Исследование, описанное в статье про Уравнения Лотки–Вольтерры, подчеркивает ее значимость в современном мире. Надеюсь, что теперь ты понял что такое Уравнения Лотки–Вольтерры и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Моделирование и Моделирование систем

Комментарии