Лекция
Привет, Вы узнаете о том , что такое транспорт через мембрану na-канал планометрическая организация na -канал объемные модели модель работы na -канала, Разберем основные их виды и особенности использования. Еще будет много подробных примеров и описаний. Для того чтобы лучше понимать что такое транспорт через мембрану na-канал планометрическая организация na -канал объемные модели модель работы na -канала , настоятельно рекомендую прочитать все из категории Физиология человека, гигиена и возрастная физиология.
При всем многообразии строения и физикохимических свойств молекул проникающих веществ можно выделить два механизма перемещения веществ через мембрану - посредством простой диффузии, т.е. без помощи специфического переносчика, и при помощи специфических переносчиков. В первом случае выделяют диффузию соединений непосредственно через липидный бислой мембраны (рис. 1-7 А) и ионов через ионные каналы (рис. 1-7 Б). Во втором случае выделяют так называемую облегченную диффузию, первично активный транспорт и, наконец, вторично активный транспорт.
Рассмотрим сначала простую диффузию. Посредством простой диффузии без помощи специального переносчика, во-первых, осуществляется транспорт соединений непосредственно через липидный бислой. В этом случае проникновение веществ в клетку идет путем их растворения в липидах клеточной мембраны, поэтому такой способ присущ водонерастворимым органическим соединениям и газам (например, кислороду и углекислому газу). Во-вторых, вещества перемещаются через ионные каналы клеточной мембраны, соединяющие цитоплазму клеток с внешней средой. Ионный канал - это белковая структура на основе мембранной α-субъединицы, образованная доменами и имеющая вид, подобный пончику с отверстием в середине - порой, через которую движутся ионы. Клетки используют этот путь для транспорта преимущественно ионов Na+, Ca2+, К+ (рис. 1-7 Б). Это пассивный ионный транспорт, который определяется градиентами концентрации и электрического поля (электрохимическим градиентом).
В данном случае понятие «градиент» отличается от его определения в математике или физике. В физико-химических или биологических системах используют термин «по градиенту», когда речь идет о движении от большего электрохимического
потенциала к меньшему, а при движении от меньшего электрохимического потенциала к большему применяют термин «против градиента».
Изменение электрохимического потенциала Δμ (без учета химической энергии, или химических потенциалов) можно записать в виде:
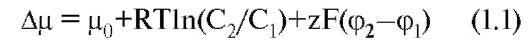
Это максимальная работа, которую можно совершить при переносе одного моля ионов. В таком случае RTln(C2/C1) равно работе по концентрированию раствора от C1 до C2, а zF(φ2-φ1) равно работе по преодолению сил электрического отталкивания, возникающих при разности потенциалов (φ2-φ1) между растворами.
При помощи специфических переносчиков осуществляется энергетически независимая облегченная диффузия ряда соединений (рис. 1-7 В, Г и Д).
Энергетически зависимый первично активный транспорт ионов Na+, Ca2+, K+ иН+ - это перенос веществ против их электрохимических градиентов с затратой энергии АТФ. Клетки в результате активного переноса ионов способны накапливать их в более высоких, по сравнению с окружающей средой, концентрациях и вопреки их заряду. Многие градиенты, возникающие на клеточной мембране и служащие необходимым условием для пассивного переноса ионов по ионным каналам, появляются именно в результате их активного транспорта. Так, градиенты концентрации K+ и Na+возникают в результате активного переноса этих ионов, т.е. работы специального Na+/К+-насоса. За счет создающейся по обе стороны мембраны разности концентраций осуществляются диффузия этих ионов по градиентам и генерация потенциалов мембраны.
Наконец, вторично активный транспорт ряда ионов и молекул тоже использует энергию, накопленную за счет потребления АТФ и затраченную на создание градиента концентрации (что дает название виду транспорта).
Рис. 1-7. Основные принципы транспорта ионов и некоторых веществ через мембрану и некоторые транспортные системы в клеточной мембране и мембране органелл.
А - продемонстрированы основные пути транспорта веществ через мембрану. Б - показаны различные типы ионных каналов, обеспечивающие транспорт ионов по электрохимическому градиенту. В - насосы, обеспечивающие транспорт ионов против градиента концентрации, обменники и транспортеры. Г и Д - транспортные системы органелл
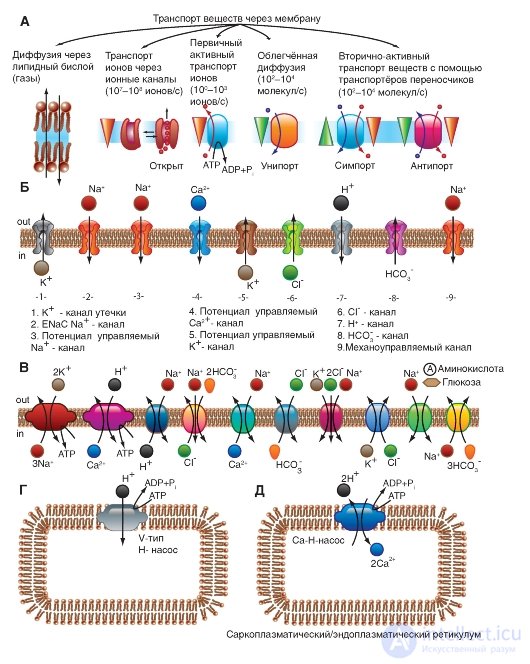
Прежде всего, рассмотрим модель молекулярной организации потенциалуправляемого Na+-канала.
Na+-канал состоит из широко разветвленной α-субъединицы, молекулярная масса которой приблизительно равна 260 кДа (рис. Об этом говорит сайт https://intellect.icu . 1-8 А). Разветвленная α-субъединица связана с добавочными β-субъединицами (рис. 1-8 Б). Na+-каналы в клетках центральной нервной системы взрослых животных содержат β1- (или β3-) и β2-субъединицы, в то время как Na+-каналы в скелетных мышцах у взрослых животных имеют только β1- субъединицу. Порообразующая α-субъединица достаточна для обеспечения функции, но кинетика и потенциалзависимость воротного механизма канала модифицируется β-субъединицами. У Na+-канала α-субъединица организована в 4 гомологичных домена (I-IV), каждый из которых содержит 6 трансмембранных α-спиралей (S1-S6- сегменты), а добавочная петля поры локализована между S5- и S6-сегментами. Петля снаружи ограничивает узкий вход в пору, в то время как S5- и S6-сегменты ограничивают внутренний, более широкий, выход из поры. Сегменты S4 в каждом домене содержат положительно заряженные аминокислотные остатки в каждой третьей позиции, выступающие в качестве воротных зарядов и двигающихся через мембрану для инициирования канальной активности в ответ на деполяризацию мембраны. Короткая внутриклеточная
петля, соединяющая гомологичные домены III и IV, выступающая в роли инактивационных ворот, образует складку, внедряющуюся внутрь канальной структуры и блокирующую пору изнутри в течение длительной деполяризации мембраны.
В последние годы для Na+-каналов была разработана новая стандартная классификация (рис. 1-8 В), основыванная на сходстве между аминокислотными последовательностями каналов. В этой классификации индивидуальный канал представлен в виде химического символа, который показывает главный проходящий ион (Na) с главным физиологическим регулятором канала (потенциалом - «voltage gate chanels»), и это записывается вместе как NaV. Число после этих символов показывает генное подсемейство (в настоящее время это только NaV1.), а число, следующее за точкой, показывает специфическую канальную изоформу (например, NaV1.1). Это последнее число было предложено для того, чтобы показать порядок, в котором был идентифицирован каждый ген. Перекрещивающиеся варианты каждого члена семейства обозначаются маленькими буквами, следующими за номером (например, NaV1.1a).
Девять изоформ Na+-каналов млекопитающих, которые были идентифицированы, функционально более чем на 50% идентичны по своей аминокислотной последовательности в трансмембранных и экстрацеллюлярных доменах, где аминокислотная последовательность одинакова. Этого достаточно для четкого выстраивания в ряд всех Na+-каналов.
Рис. 1-8. Планометрическая модель молекулярной организации потенциалуправляемого Na+-канала.
А - порообразующая α-субъединица. Основные структуры α-субъединицы показаны как трансмембранные цилиндры, представляющие собой α-спиральные сегменты. Жирные линии демонстрируют цепи полипептида каждой субъдиницы с длиной, приблизительно пропорциональной числу остатков аминокислот. α-Субъединица состоит из четырех доменов (DI-DIV), каждый из которых состоит из шести сегментов (S1-S6). Сегмент S4 - предполагаемый сенсор напряжения. Б - полная структура канала, включающая α-субъединицу и β1-, β2-субъединицы. Внеклеточные участки β1-, β2-субъединиц показаны как складки. Ψ - участки вероятного гликозилирования; P - места фосфорилирования протеинкиназой А (круги) и протеинкиназой C (ромбы); h - инактивационная частица в петле инактивационных ворот. В - подобие последовательности аминокислот и филогенетические взаимоотношения α-субъединиц потенциалуправляемых Na+-каналов. Показано сравнение аминокислотной идентичности для Na+-каналов NaV1.1- NaV1.9. Даны номер гена и номер хромосомы у человека
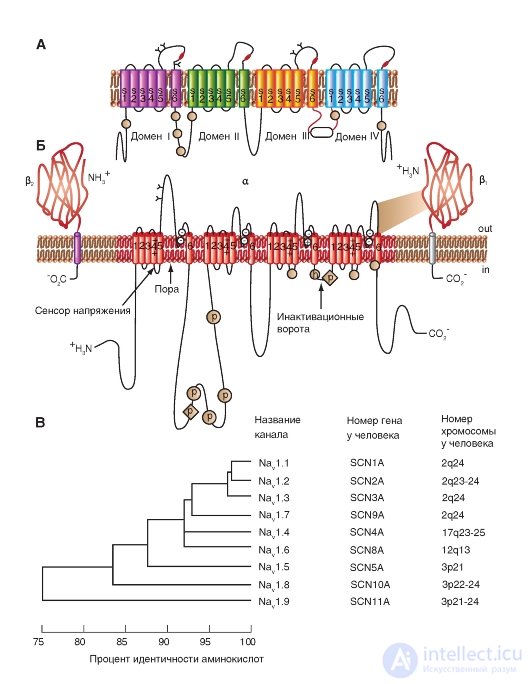
Диаметр пор Na+-канала очень маленький. Он только несколько больше, чем диаметр ионов, которые проходят через эти каналы, что предотвращает вход в них больших полярных органических молекул.
Трехмерное изображение упаковки четырех доменов порообразующей α-субъединицы потенциалуправляемого Na+-канала, каждый из которых содержит 6 пронумерованных трансмембранных сегментов (S1-S6), показано на рис. 1-9 А. Сама пора представлена полым цилиндрическим отверстием в центре упаковки. Связи между сегментами не показаны. Планометрическое изображение среза канала представлено на рис. 1-9 Б. В этом случае трехмерное изображение упаковки четырех доменов порообразую-
щей α-субъединицы потенциалуправляемого Na+-канала можно представить так, как это показано на рис. 1-9 В.
На рис. 1-9 Г канал представлен как трансмембранная макромолекула с отверстием, проходящим насквозь через центр. Ионный канал включает в себя несколько важнейших структур и в том числе устье канала, обращенное в сторону, откуда в него поступает ион (в данном случае внешняя сторона мембраны), селективный фильтр, оценивающий вид иона, активационные и инактивационные ворота, которые могут перекрывать канал для прохождения ионов, и, наконец, сенсор напряжения, управляющий работой канала. (Хотя для многих ионных каналов показано большее количество ворот канала, мы будем обсуждать их работу только с позиций активационных и инактивационных ворот.)
Рис. 1-9. Объемная модель молекулярной организации потенциалуправляемого Na+-канала.
А - структура всех четырех доменов. На схеме шесть трансмембранных сегментов каждого домена представлены в виде цилиндров и схематически объединены вместе, как это принято. Связи между сегментами и доменами не показаны. Один домен ионного канала, построенный интегральным мембранным белком, содержащим шесть трансмембранных сегментов (S1, S2, S3, S4, S5 и S6), каждый из которых имеет α-спиральную конфигурацию в пределах мембраны. Б - поперечный срез ионного канала, каждый из четырех доменов которого имеет шесть трансмембранных сегментов. В - модель объемного изображения четырех доменов с порой в середине. Г - физиологическая модель потенциалуправляемого Na+-канала
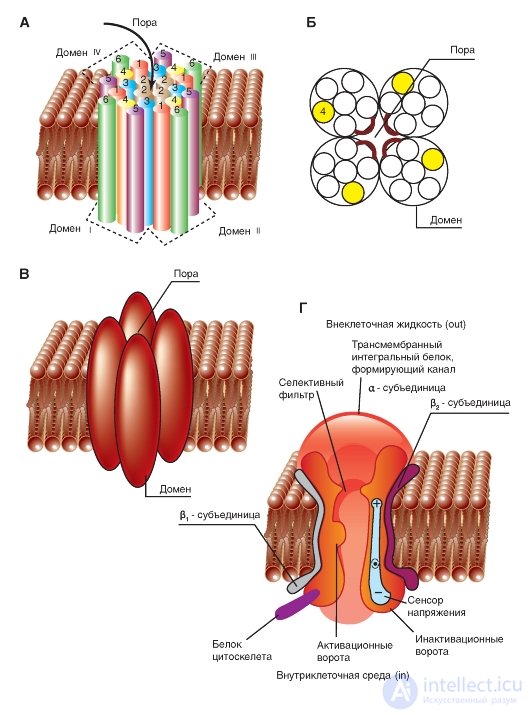
Наиболее простая и широко известная схема работы потенциалуправляемого Na+-канала показана на рис. 1-10 А. Канал представляет собой трансмембранный белок, находящийся в липидном бислое мембраны, прикрепленный к другим мембранным белкам или элементам внутриклеточного цитоскелета. Когда канал открывается, образуется водная пора, проходящая через мембрану. Устье поры намного шире, чем размер иона, только на небольшом участке в области селективного фильтра оно сужается до атомных размеров, где определяется природа иона. Гидрофильные аминокислоты формируют стенку поры, а гидрофобные аминокислоты связаны с липидным бислоем.
В настоящее время механизм работы ворот канала уже достаточно изучен. Одним из первых его попытался описать Б. Хилле. Согласно его точке зрения, основанной на экспериментах А. Ходжкина и А. Хаксли и собственных работах, в ответ на действие электрического раздражителя, т. е. на изменение трансмембранного потенциала, происходит изменение конформации белка потенциалуправляемого канала. Эти конформационные изменения регулируются электрическим полем внутри мембраны, носят стохастический характер и протекают за время от 30 мкс до 10 мс. Важно, что для открытия и закрытия канала не требуются высокоэнергетические химические соединения. Каналы открываются при одних и закрываются при других трансмембранных потенциалах. Было предположено, что электрическое поле действует на сенсор напряжения, который определяет трансмембранный потенциал. Затем сенсор напряжения должен передать эту информацию на саму канальную молекулу для ее конформационной перестройки и соответствующего изменения частоты открытия и закрытия канала.
В 1992 г. Б. Хилле предположил, что изменения конформации происходят в результате общего
перераспределения заряда в макромолекуле, образующей канал, и выражаются в виде открытия или закрытия ворот канала (рис. 1-10 А). Другими словами, возможность открытия и закрытия ворот канала контролируется сенсором напряжения. В случае потенциалуправляемых каналов сенсор должен включать много заряженных групп, которые двигаются под действием электрического поля. Необходимо заметить, что работа сенсора напряжения и ворот канала, показанная на рис. 1-10 А, представляла собой рабочую гипотезу Б. Хилле, высказанную на основании электрофизиологического изучения проводимости канала.
Рассматривая канал с позиций наличия только двух - активационных и инактивационных - ворот, можно представить следующую последовательность событий (рис. 1-10 Б). В ответ на деполяризацию мембраны происходят изменения конформации канальной молекулы: одновременно начинают смещаться активационные и инактивационные ворота, но с разной скоростью и в разных направлениях: активационные ворота стремятся открыть канал, а инактивационные - закрыть. Скорость смещения частиц, образующих активационные ворота, больше, и это приводит к открытию канала (переход из состояния покоя в состояние активации). В это время через канал проходят ионы натрия. Следующее за этим изменение трансмембранного потенциала приводит к закрытию инактивационных ворот. Это переводит канал из активированного состояния в инактивированное.
Когда канал открывается, ионный поток появляется сразу, а когда закрывается, поток ионов также сразу прекращается. На уровне одиночного канала воротные переходы стохастические; они могут быть описаны только в терминах вероятностей.
Потенциалуправляемый Na+-канал может находиться в трех состояниях: состоянии покоя, активации и инактивации, которые представлены в виде модели на рис. 1-10 Б.
Рис. 1-10. Схема работы потенциалуправляемого Na+-канала.
А - канал представляет собой трансмембранную макромолекулу с отверстием, проходящим насквозь через центр. Функциональные области ионного канала - селективный фильтр, ворота и сенсор напряжения - обнаружены в ходе электрофизиологических экспериментов. Б - модель работы потенциалуправляемого Na+-канала, имеющего активационные (или m-ворота) и инактивационные (или h-ворота). В состоянии покоя канал закрыт вследствие закрытия активационных ворот. Смещение мембранного потенциала в положительную область (до порогового значения) вызывает открытие активационных ворот. При достижении максимального для конкретной клетки потенциала канал инактивируется, т.е. происходит закрытие инактивационных ворот

Прочтение данной статьи про транспорт через мембрану na-канал планометрическая организация na -канал объемные модели модель работы na -канала позволяет сделать вывод о значимости данной информации для обеспечения качества и оптимальности процессов. Надеюсь, что теперь ты понял что такое транспорт через мембрану na-канал планометрическая организация na -канал объемные модели модель работы na -канала и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Физиология человека, гигиена и возрастная физиология
Комментарии
Оставить комментарий
Физиология человека, гигиена и возрастная физиология
Термины: Физиология человека, гигиена и возрастная физиология