Лекция
Привет, Вы узнаете о том , что такое k -канал простейшее строение потенциалуправляемые к-каналы kv семейство kv группы kir, Разберем основные их виды и особенности использования. Еще будет много подробных примеров и описаний. Для того чтобы лучше понимать что такое k -канал простейшее строение потенциалуправляемые к-каналы kv семейство kv группы kir , настоятельно рекомендую прочитать все из категории Физиология человека, гигиена и возрастная физиология.
Принятая в настоящее время классификация делит все К+-каналы на:
• потенциалуправляемые К+-каналы (KV);
• Ca2+- активируемые К+-каналы (KCa);
• K+-каналы аномального выпрямления с током входящего направления (inward rectifier K+-channels) (Kir);
• K+-каналы с двумя петлями в домене (two-P K+-channels) (К2Р).
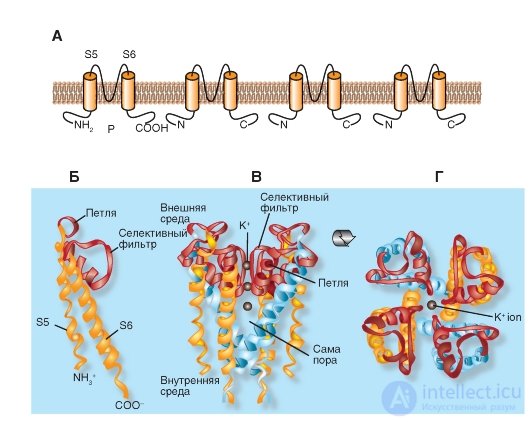
Описание молекулярной организации K+-каналов мы начнем с наиболее простой формы, представленной, например, в бактерии Streptomyces lividans. Модель молекулярной организации такого наиболее простого К+-канала бактерии Streptomyces lividans продемонстрирована на рис. 1-13.
Показанный на рис. 1-13 А К+-канал бактерий состоит из четырех одинаковых субъединиц, объединенных в единое целое тело канала.
На панелях Б, В и Г показано, что Р-петля локализована около внеклеточной поверхности мембраны и объединяет α-спирали обоих сегментов S5 и S6. Она состоит из неспиральной части, формирующей верхнюю часть поры, короткой α-спирали и удлиняющей петли, которая выдается в сужающуюся часть поры и формирует селективный фильтр. Этот фильтр позволяет проходить через канал только ионам К+, но не другим ионам. Ниже фильтра находится центральная часть поры, образованная α-спиралью сегмента S6. В тетрамерном К+-канале бактерии Streptomyces lividans сегменты S5 и S6 обозначены так, поскольку они представляют собой структурные эквиваленты сегментов S5 и S6 в потенциалуправляемом К+-канале (рис. 1-13).
На рисунке 1-13 Б показаны структурные детали канала. NH2-конец субъединицы начинается наружной спиралью (оранжевая), пронизывающей мембрану от цитоплазматической стороны до наружной поверхности. За наружной спиралью следует короткая спираль (красная), направленная в пору. Затем следует внутренняя спираль, которая возвращается к цитоплазматической стороне (оранжевая). Соединяющая петля между наружной и короткой спиралями образует структуру, формирующую наружное отверстие поры. Эта часть содержит центр связывания для неселективного блокатора К+-каналов тетраэтиламмония (ТЭА). Четыре таких петли каждой субъединицы формируют внешние стенки поры, а именно той ее части, которая ответственна за селективность канала - селективный фильтр.
Избирательность канала достигается как размером его поры, так и молекулярной организацией селективного фильтра. Диаметр селективного фильтра равен примерно 0,3 нм. Аминокислоты его стенки ориентированы так, что последовательные кольца, образованные четырьмя карбоксильными группами (по одной от каждой субъединицы), обращены внутрь поры. Диаметр поры достаточен для прохождения дегидратированного иона К+, имеющего диаметр 0,27 нм. Атомы кислорода в стенке поры заменяют гидратацию иона. Ионы меньшего размера, например Na+, с диаметром 0,19 нм, не могут проходить через K+-канал, поскольку они не могут сформировать тесную связь одновременно со всеми четырьмя кислородами стенки поры. Этот механизм в деталях представлен на рис. 1-19.
Рис. 1-13. Модель молекулярной организации наиболее простого К+-канала бактерии Streptomyces lividans.
А - этот К+-канал представляет собой тетрамер из идентичных субъединиц, каждая из которых содержит две сходные трансмембранные α-спирали, обозначаемые как S5 и S6, и короткую Р-петлю, формирующую пору. Б - организация одной из четырех субъединиц. Показаны S5 и S6 α-спирали (желтые на рисунке) и петля Р (розовая). В - тетрамерная организация канала. Вид сбоку. Г - тетрамерная организация канала. Вид сверху
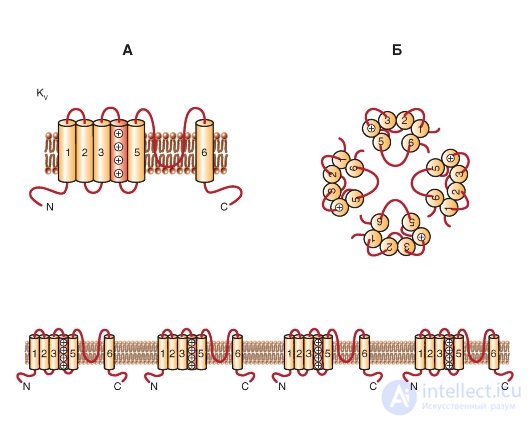
KV каналы образованы белковыми субъединицами двух типов: α-субъединицами, формирующими пору, и вспомогательными β-субъединицами. Об этом говорит сайт https://intellect.icu . Субъединицы α формируют тетрамер, чтобы образовать функциональный канал, проводящий ионы K+.
Одна субъединица ΚV-канала - белок с шестью трансмембранными сегментами (S) α-спирали (S1-S6) и расположенными в цитоплазме N- и C-терминалями. Наиболее изучены внеклеточная петля P между 5S и 6S (формирует К+-селективный фильтр и пору канала) и 4S, который положительно заряжен и формирует сенсор напряжения белка (рис. 1-14 А). Кроме того, много функций обычно приписывали большим N- и С-концевым петлям. Они включают контроль за инактивацией каналов, белок-белковое взаимодействие (по типу тетрамеризации α-субъединиц, взаимодействие с цитоплазматическими β-субъединицами и сохранение последовательностей).
KV α-субъединицы могут формировать или гомотетрамеры (homotetramers), или гетеротетрамеры (heterotetramers), вероятно, ограниченные в том же самом подсемействе. Огромное разнообразие КV-токов in vivo есть следствие существования сплайсинговых вариантов (изоформ гена, кодирующих одну и ту же α-субъединицу), сборки тетрамера α-субъединиц с различными
вспомогательными субъединицами и гетеромеризация α-субъединиц.
Как известно, ионы К+ неэквивалентно распределены между внеклеточной средой и цитоплазмой, и это создает движущую силу для выхода ионов К+ в больших физиологических пределах мембранного потенциала. КV-каналы открыты при деполяризации трансмембранного потенциала, что способствует выходу ионов К+. Деполяризация может быть представлена как аккумуляция позитивных зарядов на внешней стороне мембраны. Однако выход позитивно заряженных ионов, таких, как ионы К+, сдвигает мембранный потенциал ниже потенциала покоя. Таким образом, КV-каналы чувствуют деполяризацию и, в свою очередь, действуют так, чтобы ее устранить. Одна из наиболее охарактеризованных ролей КV-каналов заключается в том, чтобы закончить сильную деполяризацию, вызванную активацией потенциалзависимого входа катионов (совместно образующих потенциал действия). Таким образом, можно постулировать, что любые увеличения КV-канальной активности будут приводить к более эффективному окончанию деполяризации (обычно укорачивая или убирая потенциал действия). Эта функциональная способность КV-каналов также лежит в основе формирования пачечной активности (взрывной). Таким образом, активность КV-каналов играет, например, роль в кодировании информации в нервной системе.
Рис. 1-14. Планометрическая модель молекулярной организации потенциалуправляемого К+-канала (КV-канала).
А - одна субъединица КV-канала представляет собой α-спираль с шестью трансмембранными сегментами (S1-S6) и расположенными в цитоплазме N- и C-терминалями. Внеклеточная P-петля между сегментами S5 и S6 формирует селективный фильтр для ионов К+ и пору канала, а положительно заряженный сегмент S4 формирует сенсор напряжения белка. Длинные N- и C-терминальные петли контролируют инактивацию каналов и белок-белковое взаимодействие. Б - ассоциация четырех субъединиц в КV-канал
До настоящего времени в геноме человека описаны 38 генов, кодирующих различные каналы суперсемейства KV. КV-каналы включают классические категории каналов: К+-каналы задержанного выпрямления (delayed rectifier K+-channels) и быстрые транзиторные К+-каналы выходящего тока (fast transient K+-channels: fast transient K+-current IA, или transient outward current Ito).
Название «delayed rectifier K+-channels» было первоначально дано потенциалуправляемым К+- каналам гигантского аксона кальмара из-за их отсроченной активации (по сравнению с быстро активизирующимися Na+-каналами). Члены всех KVподсемейств (включая KV1-4, EAG и KCNQ) могут формировать delayed rectifier K+-channels.
Fast transient K+-channels (формирующие fast transient K+-current: IA, или transient outward current Ito) - быстрые транзиторные ^-каналы выходящего тока представляют собой каналы, активируемые низким потенциалом (low voltage-activated). Они быстро инактивирующиеся (транзиторные, иначе - временные, мимолетные K+-каналы). Каналы этого типа обычно образованы из членов
KV1 и KV4 подсемейств и вспомогательной β-субъединицы, часто необходимой для феномена быстрой инактивации. Недавно была обнаружена новая вспомогательная субъединица - CD26, родственная дипептидил аминопептидазаподобному протеину (Dipeptidyl Aminopeptidase-подобный белок, DPPX), и это позволило присвоить каналу нейронов характеристику KV4-каналов.
Филогенетические деревья KV-каналов продемонстрированы на рис. 1-15. На нем при филогенетической реконструкции, основанной на сходной аминокислотной последовательности, показаны потенциалуправляемые (voltagegated) K+-каналы семейств KV1-KV6, KV7 и KV8-KV9. Среди этой группы только KV1.8 в настоящее время испытывает недостаток в названии HGNC. Пять членов группы KV семейства KV7 (KCNQ1-KCNQ5) в настоящее время (с 2005 г.) могут быть внесены в союз с другими белками канала KV, и поэтому не показаны в отдельном филогенетическом дереве, как это представлялось ранее. Три оставшиеся семейства KV - KV10, KV11 и KV12 - достаточно близко связаны друг с другом, чтобы быть показанными в отдельном филогенетическом дереве.
Рис. 1-15. Филогенетические деревья для семейства KV.
А - филогенетическое дерево для семейства КV1-К^9-каналов. Аминокислотные последовательности KV-канала включают KV1-KV6 и KV8-KV9 семейства, которые были внесены до 2005 г. Б - последовательности KV7.1-KV7.5, KV6.4 и KV8.2 были добавлены к существующему филогенетическому дереву. Только гидрофобные ядра канала (S1-S6) использовались для анализа. В - филогенетическое дерево для семейств KV10-KV12-каналов

Следующий класс 2S-белков включает три группы каналов inward rectifiers channels (Kir), которые можно назвать, хотя и очень длинно, как «К+-каналы аномального выпрямления с током входящего направления». N- и С-терминали этих каналов также расположены в цитоплазме, P-петля между двумя трансмембранными доменами формирует пору, и функционирующий канал представляет собой тетрамер этих 2S/1P-субъединиц.
Клонирование первых inward rectifiers K+-channels - Kir1.1 (ROMK1) и Kir2.1 (IRK1) - было осуществлено в 1993 г., и последовательность новых членов этого семейства была идентифицирована, включая К-канал, связанный с G-белком, (Kir3) и АТФ-чувствительный К-канал (Kir6). Эти каналы играют важную физиологическую роль в функции многих органов, включая мозг, сердце, почки, эндокринные клетки, слуховые клетки и клетки сетчатки. Молекулярная организация этих каналов приведена на рис. 1-16 А. Филогенетическое дерево, показанное на рис. 1-16 Б, иллюстрирует родственные связи, основанные на последовательности аминокислотных остатков, между семью основными
подсемействами Kir. В настоящее время новые члены этого семейства не были идентифицированы. Поскольку это филогенетическое дерево появилось только в 2002 г. в издании IUPHAR, маловероятно, что какие-либо другие новые члены были обнаружены за это время.
В последнее время метод рентгеноструктурной кристаллографии дал возможность описать структуру цитоплазматического региона Kir3.1, полную структуру бактериальных Kir1.1 и цитоплазматическую область Kir2.1. Эти данные демонстрируют, что inward rectifiers K+-channels имеют длинную пору, уходящую в цитоплазму, и подчеркивают важность отрицательных зарядов аминокислотных остатков на стенке цитоплазматической части поры, которая играет важную роль аномального выпрямления. Эти исследования обеспечили понимание механизмов регулирования ворот канала посредством лигандов через G-белки и фосфатидилинозитол 4,5-бифос- фат. Информация, полученная благодаря анализу кристаллической структуры, чрезвычайно ценна, так как позволяет оценить структурно-функциональные отношения. Также примечательны издания, посвященные изучению динамических аспектов функции канала.
Рис. 1-16. Молекулярная организация inwardly rectifying potassium channels - K+-каналов аномального выпрямления с током входящего направления (Kir).
А - одна субъединица К1r-канала представляет собой α-спираль с двумя трансмембранными сегментами (S5 и S6), расположенными в цитоплазме N- и C-терминалями, и короткую Р-петлю, формирующую пору. Б - ассоциация четырех субъединиц в Кir-канал. В - филогенетическое дерево для Кir-каналов

Прочтение данной статьи про k -канал простейшее строение потенциалуправляемые к-каналы kv семейство kv группы kir позволяет сделать вывод о значимости данной информации для обеспечения качества и оптимальности процессов. Надеюсь, что теперь ты понял что такое k -канал простейшее строение потенциалуправляемые к-каналы kv семейство kv группы kir и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Физиология человека, гигиена и возрастная физиология

Комментарии