Лекция
Привет, Вы узнаете о том , что такое са-канал планометрическая организация кальциевые токи молекулярная организация кальциевого канала, Разберем основные их виды и особенности использования. Еще будет много подробных примеров и описаний. Для того чтобы лучше понимать что такое са-канал планометрическая организация кальциевые токи молекулярная организация кальциевого канала , настоятельно рекомендую прочитать все из категории Физиология человека, гигиена и возрастная физиология.
Потенциалуправляемые Са2+-каналы позволяют ионам Са2+ входить в клетки, когда мембрана деполяризована. Роль Са2+-каналов особенно значительна для взаимосвязи электрических потенциалов, возникающих на мембране клетки, с физиологическими процессами, происходящими внутри клетки. Повышение цитозольной концентрации Ca2+ вызывает различные клеточные реакции, например сокращение, секрецию, выброс медиатора, процессы транскрипции.
Са2+-каналы представляют собой комплекс белков, образованный из α1-субъединицы и вспомогательных α2δ-, β- и γ-субъединиц. Субъединица α1 формирует проводящую пору, она содержит сенсор напряжения и аппарат ворот канала. Петля между трансмембранными сегментами S5 и S6 в каждом домене определяет селективность и проводимость канала. Селективный фильтр Са2+-канала должен узнать ион Са2+ уже на входе в канал. Эти события достаточно редки по сравнению с входами ионов Na+, количество которых во внеклеточной среде примерно в 100 раз больше. Хотя ионы Са2+ и Na+ имеют идентичный диаметр (2 Ао), канал может выбирать Са2+ в большей степени, чем Na+, в соотношении 1000:1. Никакое сито не может так эффективно дифференцировать ионы идентичного размера. Скорее всего, пора содержит специфическое место,
которое способно связывать Са2+ при его очень низких концентрациях в растворе (10-6 M), а все другие физиологические ионы связывает в намного более высоких концентрациях.
Са2+-каналы были классифицированы на основе двух принципов:
• химический символ главного иона, для которого они проницаемы (в данном случае ионы
Ca2+);
• принцип физиологической регуляции работы канала, имея в виду, что эти каналы потенциалуправляемые (voltage gated calcium channels) Са2+-каналы, что позволило ввести обозначение СаV (рис. 1-11 В).
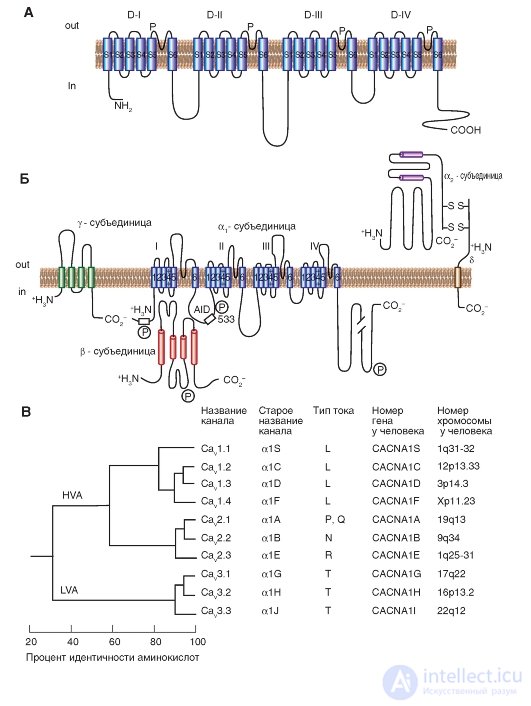
Первая цифра после обозначения соответствует в субъединице номеру генного подсемейства (от 1 до 3 в настоящее время), а вторая цифра, которая ставится после точки, соответствует порядку открытия оригинальной изоформы α1субъединицы внутри этого подсемейства (от 1 до m). Соответственно этой номенклатуре, СаV1 подсемейство (от СаV1.1 до СаV1.4) включает каналы, содержащие α1S, α1C, α1D и α1F, для которых характерен L-тип кальциевых токов. Подсемейство СаV2 (от СаV2.1 до СаV2.3) включает каналы, содержащие α1Α, α1Β и α1Ε, для которых характерен P/Q-тип, N-тип и R-тип кальциевых токов соответственно. Подсемейство СаV3 (от СаV3.1 до СаV3.3) включает каналы, содержащие α1G, α1Η и α1I, для которых характерен T-тип кальциевых токов.
Рис. Об этом говорит сайт https://intellect.icu . 1-11. Планометрическая модель молекулярной организации потенциалуправляемого Ca2+-канала.
А - основная α1-субъединица, формирующая пору. Основные структуры α1-субъединицы показаны как трансмембранные цилиндры, представляющие собой α-спиральные сегменты. Б - модель выполнена на основе экспериментальных данных изучения структуры Са2+-каналов, состоящих из пяти субъединиц. В дополнение к α1-субъединице представлена локализованная внутриклеточно β-субъединица и расположенная внеклеточно α2-субъединица, соединенная дисульфидной связью с δ-субъединицей в комплекс α2δ. Масштаб длины линий приблизительно соответствует длине полипептидных сегментов. Участки различий в α1-субъединице между некоторыми Са2+-каналами мембран клеток расположены в NH2-концевой части в гидрофобных сегментах DIS6 и DIVS3 и в линкере (цитоплазматическом связующем участке) между доменами I и II. AID (α1-interaction domain) - первичная область связывания для всех CaVα1-субъединиц с CaVβ-субъединицами, названная как α1-взаимодействующий домен. Символом «Р» обозначены места фосфорилирования разными протеинкиназами. В - последовательное сходство α1-субъединиц потенциалуправляемых Са2+-каналов. Продемонстрировано филогенетическое дерево первичных последовательностей Са2+-каналов. В результате сравнения пар последовательностей четко дифференцированы три семейства с межсемейственными последовательностями, идентифицированными более чем 80% (CaV1, CaV2, CaV3). Далее сходство последовательности было определено для каждого семейства, и эти три последовательности были сравнены одна с другой с межсемейственными последовательностями, идентифицированными на 52% (CaV1 по сравнению с CaV2) и на 28% (CaV3 по сравнению с CaV1 или с CaV2)
Са2+-каналы были обнаружены практически во всех клетках. Са2+-токи, зарегистрированные в различных типах клеток, имеют определенные физиологические и фармакологические свойства. Изначальная буквенная номенклатура была предложена исходя из кинетик Са2+-токов. L-тип (от «long-lasting» - долго длящийся) Са2+-токов требует сильной деполяризации для активации, он долго длящийся и блокируется органическими антагонистами L-типа Са2+-каналов, включая дигидропиридины, фенилалкиламины и бензотиазепины. Са2+-токи L-типа являются главными в мышцах и эндокринных клетках, где они инициируют сокращение и секрецию. N-тип (от «neither long nor transient» - ни L, ни T), P/Q-тип и R-тип кальциевых токов также требуют сильной деполяризации для активации. Они относительно нечувствительны к антагонистами L-типа Са2+-каналов, но блокируются специфическими полипептидными токсинами из ядов улитки или паука. Эти токи в основном выражены в нейронах, где они инициируют нейротрансмиссию для большинства быстрых синапсов и также опосредуют вход Са2+ в клеточные тела и дендриты. T-тип (от «transient» - преходящий) Са2+-токов активируется слабой деполяризацией, и эти токи мимолетные (преходящие). Они нечувствительны к органическим антагонистам и токсинам змей и пауков, которые используются для определения N- и P/Q Са2+-токов. Са2+-токи T-типа выражены в большом спектре клеточных типов, где они вовлечены в развитие потенциала действия и важны в клетках и тканях, обладающих ритмической активностью.
Са2+-каналы, которые были охарактеризованы биохимическими методами, представляют собой комплекс белков, образованный из четырех или пяти определенных субъединиц, кодирующихся большим семейством генов.
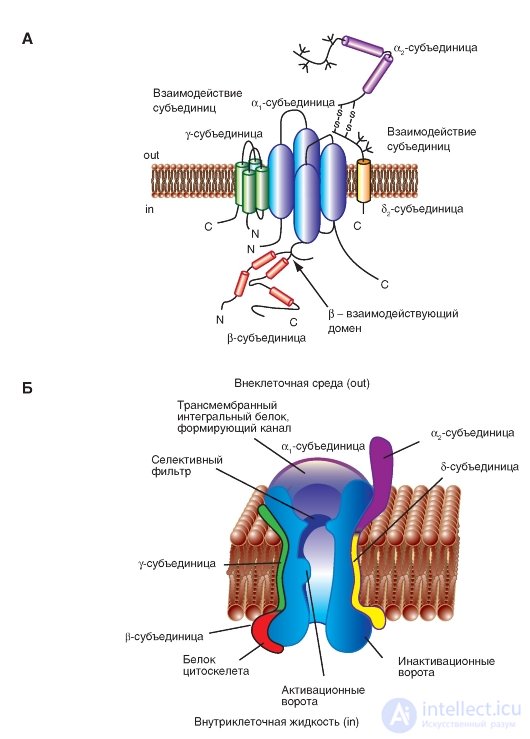
На молекулярном уровне Са2+-канал составлен из формирующей пору трансмембранной α1-субъединицы и вспомогательных α2δ, β и γ-субъединиц (рис. 1-12 А). При этом α2-субъединица находится на внешней стороне мембраны, β-субъединица - на внутренней стороне мембраны, а δ- и γ-субъединицы представляют собой трансмембранные структуры. Структура Са2+-канала сердечной и гладкой мышцы - это структура с четырьмя субъединицами канала, и она напоминает Са2+-каналы нейрона. Вспомогательные α2δ- и β-субъединицы увеличивают транспорт ионов Са2+ через пору, образованную α1-субъединицей, и модулируют потенциалзависимую кинетику канала. Внутриклеточная β-субъединица и трансмембранный дисульфид-связанный комплекс α2δ-субъединиц являются компонентами большинства типов Са2+-каналов. В дополнение к роли β-субъединицы в транспорте ионов через канал, она также играет принципиальную роль в электромеханическом сопряжении в мышечных волокнах скелетных мышц. Взаимодействия субъединиц α2δ- и β-субъединицы в настоящее время хорошо изучены. Наличие субъединицы γ было также обнаружено в Са2+-каналах скелетных мышц, в кардиомиоцитах и нейронах головного мозга. Хотя эти добавочные субъединицы модулируют свойства ионного канала, фармакологические и электрофизиологические особенности Са2+-каналов в основном связаны с существованием α1-субъединиц.
β-субъединицы Са2+-каналов способны формировать гетерогенные комплексы in vivo и in vitro. β-субъединицы и α1-субъединица обладают консервативными областями взаимодействия, что способствует формированию гетерогенных комплексов канала. α2δ-субъединицы модулируют различные α1-субъединицы in vitro, и возможно, что эта гетерогенность распространяется на ситуации in vivo. Такое разнообразие взаимодействия субъединиц потенциально увеличило бы число возможных вариантов каналов с отличающимися биофизическими и физиологическими свойствами, что может обеспечивать разнообразие клеточных ответов.
Физиологическая модель потенциалуправляемого Na+-канала показана на рисунке (рис. 1-12 Б).
Рис. 1-12. Объемная модель взаимодействий субъединиц потенциалуправляемого Ca2+-канала.
А - γ-субъединица изображена как трансмембранный белок с четырьмя сегментами с внутриклеточным N- и C-концами. Первая половина γ-субъединицы взаимодействует с α1-субъединицей. Первая внеклеточная петля содержит заряженные остатки и участки гликозилирования. Определены взаимодействующие участки α2δ- и β-субъединица (β-взаимодействующий домен) с α1-субъединицей. Б - физиологическая модель потенциалуправляемого Na+-анала. Показана основная α1-субъединица, формирующая пору и дополнительные α2-, δ-, γ- и β-субъединицы
Прочтение данной статьи про са-канал планометрическая организация кальциевые токи молекулярная организация кальциевого канала позволяет сделать вывод о значимости данной информации для обеспечения качества и оптимальности процессов. Надеюсь, что теперь ты понял что такое са-канал планометрическая организация кальциевые токи молекулярная организация кальциевого канала и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Физиология человека, гигиена и возрастная физиология
Комментарии
Оставить комментарий
Физиология человека, гигиена и возрастная физиология
Термины: Физиология человека, гигиена и возрастная физиология