Лекция
Привет, сегодня поговорим про биологический нейрон, обещаю рассказать все что знаю. Для того чтобы лучше понимать что такое биологический нейрон, его кибернетическая модель , настоятельно рекомендую прочитать все из категории Computational Neuroscience (вычислительная нейронаука) Теория и приложения искусственных нейронных сетей.
Нейробиология. биологический нейрон , его строение и функции. Об'единение нейронов в сети. Биологическая изменчивость и обучаемость нейронных сетей. Кибернетическая модель нейрона - формальный нейрон МакКаллока и Питса. Обучение нейрона задаче детектирования границы яркости.
Эта лекция посвящена биологическим основаниям науки о вычислительных нейронных сетях. Также, как и в предыдущей лекции, изложение будет носить справочный характер, и предназначено для читателя, не имеющего специальных знаний по биологии. Более глубокие профессиональные сведения можно найти в относительно недавно переведенной книге Н.Грина, У. Стаута и Д.Тейлора, а также в монографии Г.Шеперда. Для ознакомительного чтения можно порекомендовать книгу Ф.Блума, А.Лейзерсона и Л.Хофстедтера.
На протяжении книги нашей основной целью будет исследование методов и кибернетических систем, имитирующих функции мозга при решениии информационных задач. Такой путь разработки искусственных вычислительных систем во многом представляется естественным - высшие биологические организмы, и особенно человек, легко справляются с такими, крайне сложными при математическом рассмотрении проблемами, как, например, распознавание образов (зрительных, слуховых, сенсорных и других), память и устойчивое управление движением тела. Биологический фундамент при изучении этих функций является крайне важным, природное многобразие дает исключительно богатый исходный материал для направленного создания искусственных моделей.
В завершении лекции будет представлена классическая кибернетическая модель нейрона - так называемый формальный нейрон Маккалока и Питтса. Некоторые свойства формального нейрона будут изучены на задаче детектирования границы перехода "черное-белое" в простом изображении.
К предмету нейробилологии относится изучение нервной системы и ее главного органа - мозга. Принципиальным вопросом для этой науки является выяснение соотношения междустроением нервной системы и ее функцией. При этом рассмотрение проводится на на нескольких уровнях: молекулярном, клеточном, на уровне отдельного органа, организма в целом, и далее на уровне социальной группы. Таким образом, классический нейробилогический подход состоит в последовательном продвижении от элементарных форм в направлении их усложнения.
Для наших практических целей отправной точкой будет клеточный уровень. По современным представлениям, именно на нем совокупность элементарных молекулярных химико-биологических процессов, протекающих в отдельной клетке, формирует ее как элементарных процессор, способный к простейшей переработке информации.
Элементом клеточной структуры мозга является нервная клетка - нейрон. Нейрон в своем строении имеет много общих черт с другими клетками биоткани: тело нейрона окружено плазматической мембраной, внутри которой находится цитоплазма, ядро и другие составляющие клетки. Однако нервная клетка существенно отличается от иных по своемуфункциональному назначению. Нейрон выполняет прием, элементарное преобразование и дальнейшую передачу информации другим нейронам. Информация переносится в виде импульсов нервной активности, имеющих электрохимическую природу.
Нейроны крайне разнообразны по форме, которая зависит от их местонахождения в нервной системе и особенностей фунционирования. На Рис. 3.1. приведена схема строения "типичного" нейрона. Тело клетки содержит множество ветвящихся отростков двух типов. Отростки первого типа, называемые дендритами за их сходство с кроной раскидистого дерева, служат в качестве входных каналов для нервных импульсов от других нейронов. Эти импульсы поступают в сому или тело клетки размером от 3 до 100 микрон, вызывая ее специфическое возбуждение, которое затем распространяется по выводному отростку второго типа - аксону. Длина аксонов обычно заметно превосходит размеры дентритов, в отдельных случаях достигая десятков сантиметров и даже метров. Гигантский аксон кальмара имеет толщину около миллиметра, и именно наблюдение за ним послужило выяснению механизма передачи нервных импульсов между нейронами.
Тело нейрона, заполненное проводящим ионным раствором, окружено мембраной толщиной около 75 ангстрем, обладающей низкой проводимостью. Между внутреннней поверхностью мембраны аксона и внешней средой поддерживается разность электрических потенциалов. Это осуществляется при помощи молекулярного механизма ионных насосов, создающих различную концентрацию положительных ионов K+ и Na+ внутри и вне клетки. Проницаемость мембраны нейрона селективна для этих ионов. Внутри аксона клетки, находящейся в состоянии покоя, активный транспорт ионов стремится поддерживать концентрацию ионов калия более высокой, чем ионов натрия, тогда как в жидкости, окружающей аксон, выше оказывается концентрация ионов Na+. Пассивная диффузия более подвижных ионов калия приводит к их интенсивному выходу из клетки, что обуславливает ее общий отрицательный относительно внешней среды потенциал покоя, составляющий около -65 милливольт.

Рис. 3.1.Общая схема строения биологического нейрона.
Под воздействием стимулирующих сигналов от других нейронов мембрана аксона динамически изменяет свою проводимость. Это происходит, когда суммарный внутренний потенциал превышает пороговое значение масштаба -50 мв. Мембрана на короткое время, сотавляющее около 2 миллисекунд, изменяет свою полярность (деполяризуется) и достигает потенциала действия около +40 мв. На микроуровне это объясняется кратковременным повышением проницаемости мембраны для ионов Na+ и активным поступлением их в аксон. В дальнейшем, по мере выхода ионов калия, положительный заряд с внутренней стороны мембраны меняется на отрицательный, и наступает так называемый период рефрактерности, длящийся около 200 мс. В течении этого времени нейрон является полностью пассивным, практически неизменно сохраняя потенциал внутри аксона на уровне около -70 мв.
Импульс деполяризации клеточной мембраны, называемый спайком, распространяется вдоль аксона практически без затухания, поддерживаясь локальными ионными градиентами. Скорость перемещения спайка является относительно невысокой и составляет от 100 до 1000 сантиметров в секунду.
Возбуждение нейрона в виде спайка передается другим нейронам, которые таким образом объеденены в проводящую нервные импульсы сеть. Об этом говорит сайт https://intellect.icu . Участки мембраны на аксоне, где размещаются области контакта аксона данного нейрона с дендритами другими нейронов, называются синапсами. В области синапса, имеющего сложное строение, происходит обмен информацией о возбуждении между нейронами. Механизмы синаптической передачи достаточно сложны и разнообразны. Они могут иметь химическую и электрическую природу. В химическом синапсе в передаче импульсов участвуют специфические химические вещества - нейромедиаторы, вызывающие изменения проницаемости локального участка мембраны. В зависимости от типа вырабатываемого медиатора синапс может обладать возбуждающим (эффективно проводящим возбуждение) или тормозящим действием. Обычно на всех отростках одного нейрона вырабатывается один и тот же медиатор, и поэтому нейрон в целом функционально является тормозящим или возбуждающим. Это важное наблюдение о наличии нейронов различных типов в последующих главах будет существенно использоваться при проектировании искуственных систем.
Взаимодействующие между собой посредством передачи через отростки возбуждений нейроны формируют нейронные сети. Переход от рассмотрения отдельного нейрона к изучению нейронных сетей является естественным шагом в нейробиологической иерархии.
Общее число нейронов в центральной нервной системе человека достигает 1010 - 1011, при этом каждая нервная клетка связана в среднем с 103 - 104 других нейронов. Установлено, что в головном мозге совокупность нейронов в объеме масштаба 1 мм3формирует относительно независимую локальную сеть, несущую определенную функциональную нагрузку.
Выделяют несколько (обычно три) основных типов нейронных сетей, отличающихся структурой и назначением. Первый тип составляют иерархические сети, часто встречающиеся в сенсорных и двигательных путях. Информация в таких сетях передается в процессе последовательного перехода от одного уровня иерархии к другому.

Рис. 3.2.Структура простой рефлекторной нейронной сети.
Нейроны образуют два характерных типа соединений - конвергентные, когда большое число нейронов одного уровня контактирует с меньшим числом нейронов следующего уровня, и дивергентные, в которых контакты устанавливаются со все большим числом клеток последующих слоев иерархии. Сочетание конвергентных и дивергентных соединений обеспечивает многократное дублирование информационных путей, что является решающим фактором надежности нейронной сети. При гибели части клеток, сохранившиеся нейроны оказываются в состоянии поддерживать функционирование сети. Ко второму типу нейронных сетей относятся локальные сети, формируемые нейронами с ограниченными сферами влияния. Нейроны локальных сетей производят переработку информации в пределах одного уровня иерархии. При этом функционально локальная сеть представляет собой относительно изолированную тормозящую или возбуждающую структуру. Важную роль также играют так называемые дивергентные сети с одним входом. Командный нейрон, находящийся в основании такой сети может оказывать влияние сразу на множество нейронов, и поэтому сети с одним входом выступают согласующим элементом в сложном сочетании нейросетевых систем всех типов.
Рассмотрим схематически нейронную сеть, формирующую простой рефлекторную цепь с передачей возбуждения от раздражителя к двигательной мышце (Рис. 3.2).
Сигнал внешнего раздражителя воспринимается сенсорными нейронами, связанными с чувствительными клетками-рецепторами. Сенсорные нейроны формируют первый (нижний) уровень иерархии. Выработанные ими сигналы передаются нейронам локальной сети, содержащим множество прямых и обратных связей с сочетанием дивергентных и конвергентных соединений. Характер преобразованного в локальных сетях сигнала определяет состояние возбуждения моторных нейронов. Эти нейроны, составляющие верхний в рассматриваемой сети уровень иерархии, образно говоря, "принимают решение", которое выражается в воздействии на клетки мышечной ткани посредством нервно-мышечных соединений.
Структура основных типов нейронных сетей генетически предопределена. При этом исследования в области сравнительной нейроанатомии говорят о том, что по фундаментальному плану строения мозг очень мало изменился в процессе эволюции. Однако детерминированные нейронные структуры демонстрируют свойства изменчивости, обуславливающие их адаптацию к конкретным условиям функционирования.
Генетическая предопределенность имеет место также и в отношении свойств отдельных нейронов, таких, например, как тип используемого нейромедиатора, форма и размер клетки. Изменчивость на клеточном уровне проявляется в пластичности синаптических контактов. Характер метаболической активности нейрона и свойства проницаемости синаптической мемраны могут меняться в ответ на длительную активизацию или торможение нейрона. Синаптический контакт "тренируется" в ответ на условия функционирования.
Изменчивость на уровне сети связана со спецификой нейронов. Нервная ткань практически лишена характерной для других типов тканей способности к регенерации путем деления клеток. Однако нейроны демонстрируют способность к формированию новых отростков и новых синаптических контактов. Ряд экспериментов с преднамеренным повреждением нервных путей указывает, что развитие нейронных ответвлений сопровождается конкуренцией за обладание синаптическими участками. Это свойство в целом обеспечивает устойчивость функционирования нейронный сетей при относительной ненадежности их отдельных компонент - нейронов.
Специфическая изменчивость нейронных сетей и свойств отдельных нейронов лежит в основе их способности к обучению - адаптации к условиям функционирования - при неизменности в целом их морфологической структуры. Следует заметить, однако, что рассмотрение изменчивости и обучаемости малых групп нейронов не позволяет в целом ответить на вопросы об обучаемости на уровне высших форм психической деятельности, связанных с интеллектом, абстрактным мышлением, речью.
Прежде чем перейти к рассмотрению моделей нейронов и искусственных нейронных сетей, сформулируем общие фактологические положения о биологических нейронных сетях. Здесь мы полностью следуем изложенному в книге Ф.Блума, А.Лейзерсона и Л.Хофстедтера.
Основными действующими элементами нервной системы являются отдельные клетки, называемые нейронами. Они имеют ряд общих с клетками других типов черт, при этом сильно отличаясь от них по своей конфигурации и функциональному назначению. Активность нейронов при передаче и обработке нервных импульсов регулируется свойствами мембраны, которые могут меняться под воздействием синаптических медиаторов. Биологические функции нейрона могут меняться и адаптироваться к условиям функционирования. Нейроны объединяются в нейронные сети, основные типы которых, а также схемы проводящих путей мозга являются генетически запрограммированными. В процессе развития возможно локальное видоизменение нейронных сетей с формированием новых соединений между нейронами. Отметим также, что нервная система содержит помимо нейронов клетки других типов.
Исторически первой работой, заложившей теоретический фундамент для создания искусственных моделей нейронов и нейронных сетей, принято считать опубликованную в 1943 г. статью Уоррена С.Мак-каллока и Вальтера Питтса "Логическое исчислени идей, относящихся к нервной активности". Главный принцип теории Маккалока и Питтса заключается в том, что произвольные явления, относящиеся к высшей нервной деятельности, могут быть проанализированы и поняты, как некоторая активность в сети, состоящей из логических элементов, принимающих только два состояния ("все или ничего"). При этом для всякого логического выражения, удовлетворяющего указанным авторами условиям, может быть найдена сеть логических элементов, имеющая описываемое этим выражением поведение. Дискуссионные вопросы, касающиеся возможности моделирования психики, сознания и т.п. находятся за рамками этой книги.
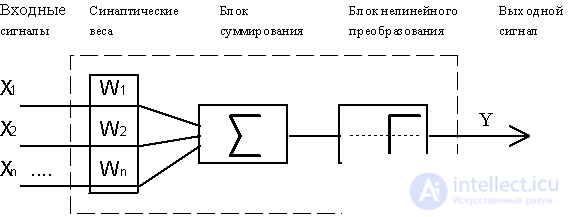
Рис.4.1. Функциональная схема формального нейрона Маккалока и Пиитса.
В качестве модели такого логического элемента, получившего в дальнейшем название "формальный нейрон", была предложена схема, приведенная на Рис. 4.1. С современной точки зрения, формальный нейрон представляет собой математическую модель простого процессора, имеющего несколько входов и один выход. Вектор входных сигналов (поступающих через "дендриды") преобразуется нейроном в выходной сигнал (распространяющийся по "аксону") с использованием трех функциональных блоков: локальной памяти, блока суммирования и блока нелинейного преобразования.
Вектор локальной памяти содержит информацию о весовых множителях, с которыми входные сигналы будут интерпретироваться нейроном. Эти переменные веса являются аналогом чувствительности пластических синаптических контактов. Выбором весов достигается та или иная интегральная функция нейрона.
В блоке суммирования происходит накопление общего входного сигнала (обычно обозначаемого символом net), равного взвешенной сумме входов:
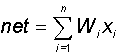
В модели Маккалока и Питтса отсутствуют временные задержки входных сигналов, поэтому значение net определяет полное внешненее возбуждение, воспринятое нейроном. Отклик нейрон далее описывается по принципу "все или ничего", т. е. переменная подвергается нелинейному пороговому преобразованию, при котором выход (состояние активации нейрона) Y устанавливается равным единице, если net > Q, и Y=0 в обратном случае. Значение порога Q (часто полагаемое равным нулю) также хранится в локальной памяти.
Фомальные нейроны могут быть объединены в сети путем замыкания выходов одних нейронов на входы других, и по мысли авторов модели, такая кибернетическая система с надлежаще выбранными весами может представлять произвольную логическую функцию. Для теоретического описания получаемых нейронных сетей предлагался математический язык исчисления логических предикатов.
Нужно отметить, что сегодня, спустя 50 лет после работы Маккалока и Питтса, исчерпывающей теории синтеза логических нейронных сетей с произвольной функцией, по-видимому, нет. Наиболее продвинутыми оказались исследования в области многослойных систем и сетей с симметричными связями. Большинство моделей опираются в своей основе на различных модификациях формального нейрона. Важным развитием теории формального нейрона является переход к аналоговым (непрерывным) сигналам, а также к различным типам нелинейных переходных функций. Опишем наиболее широко используемые типы переходных функций Y=f(net).


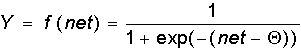
Как указывалось еще С.Гроссбергом, сигмоидальная функция обладает избирательной чувствительностью к сигналам разной интенсивности, что соответсвует биологическим данным. Наибольшая чувствительность наблюдается вблизи порога, где малые изменения сигнала net приводят к ощутимым изменениям выхода. Напротив, к вариациям сигнала в областях значительно выше или ниже порогового уровня сигмоидальная функция не чувствительна, так как ее производная при больших и малых аргументах стремится к нулю.
В последнее время также рассматриваются математические модели формальных нейронов, учитывающие нелинейные корреляции между входами. Для нейронов Маккалока и Питтса предложены электротехнические аналоги, позволяющие проводить прямое аппаратное моделирование.
Способность формального нейрона к обучению проявляется в возможности изменения значений вектора весов W, соответствующей пластичности синапсов биологических нейронов. Рассмотрим обучение формального нейрона на примере простейшей задачи детектирования границы. Пусть имеется образ, составленный из одномерной цепочки черных и белых клеток. Зачерненные клетки соответсвуют единичному сигналу, а белые клетки - нулевому. Сигнал на входах формального нейрона устанавливается равным значениям пар примыкающих клеток рассматриваемого образа. Нейрон обучается всякий раз возбуждаться и выдавать единичный выходной сигнал, если его первый вход (на Рис. 4.2. - левый) соединен с белой клеткой, а второй (правый) - с черной. Таким образом, нейрон должет служить детектором границы перехода от светлого к темному тону образа.
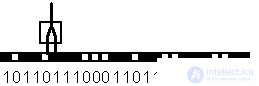
Рис. 4.2. Формальный нейрон с двумя входами, занятый обработкой образа в виде одномерной цепочки черных и белых клеток.
Функция, выполняемая нейроном, определяется следующей таблицей.
|
Вход 1 |
Вход 2 |
Требуемый выход |
|
1 |
1 |
0 |
|
1 |
0 |
0 |
|
0 |
1 |
1 |
|
0 |
0 |
0 |
Для данной задачи значения весов и порога нейрона могут быть предъявлены и без специальной процедуры обучения. Легко убедиться, что нужным требованиям удовлетворяет набор Q = 0, W1 = -1, W1 = +1. В случае задачи детектирования границы перехода от темного к светлому веса нужно поменять местами.
В общем случае для подстройки весов при обучении нейрона разработаны различные алгоритмы, которые будут рассматриваться в применении к конкретным типам нейронных сетей, составленных из формальных нейронов.
В общем, мой друг ты одолел чтение этой статьи об биологический нейрон. Работы впереди у тебя будет много. Смело пиши комментарии, развивайся и счастье окажется в твоих руках. Надеюсь, что теперь ты понял что такое биологический нейрон, его кибернетическая модель и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Computational Neuroscience (вычислительная нейронаука) Теория и приложения искусственных нейронных сетей
Ответы на вопросы для самопроверки пишите в комментариях, мы проверим, или же задавайте свой вопрос по данной теме.

Комментарии