Лекция
Привет, мой друг, тебе интересно узнать все про развитие психической деятельности в пренатальном периоде, тогда с вдохновением прочти до конца. Для того чтобы лучше понимать что такое развитие психической деятельности в пренатальном периоде , настоятельно рекомендую прочитать все из категории Сравнительная психология и Зоопсихология.
Поведение эмбриона является во многих отношениях основой всего процесса развития поведения в онтогенезе. Как у беспозвоночных, так и у позвоночных установлено, что развивающийся организм производит еще в пренатальном (дородовом) периоде движения, которые являются элементами будущих двигательных актов, но еще лишены соответствующего функционального значения, т. е. еще не могут играть приспособительную роль во взаимодействии животного со средой его обитания. Эта функция появляется лишь в постнатальном периоде его жизни. В таком смысле можно говорить о преадаптационном значении эмбрионального поведения.
Академик И. И. Шмальгаузен выделил несколько типов онтогенетических корреляций, среди которых особый интерес для познания закономерностей развития поведения представляют «эргонтические корреляции» (или функциональные в узком смысле), т. е. соотношения между частями и органами, обусловленные функциональными зависимостями между ними. Шмальгаузен при этом имел в виду типичные («окончательные») функции органов. Таковыми являются уже на ранних стадиях эмбриогенеза, например, функции сердца или почек зародыша. В качестве характерного примера эргонтических корреляций Шмальгаузен указывает и на зависимость между развитием нервных центров и нервов и развитием периферических органов — органов чувств и конечностей. При экспериментальном удалении этих органов соответствующие элементы нервной системы недоразвиваются.
Как установил еще в 30-е годы Б. С. Матвеев, эмбриогенез характеризуется коррелятивными сдвигами в соотношениях между развивающимися органами, которые являются следствием нарушения взаимосвязей частей зародыша в развивающемся организме. В результате возникают функциональные изменения, определяющие характер деятельности формирующихся органов.
Эти выводы ведущих советских зоологов показывают всю сложность морфофункциональных связей и взаимозависимостей, определяющих формирование поведения животных в эмбриогенезе. Это нашло свое отражение и в концепции системогенеза П. К. Анохина. «Развитие функции, — говорил Анохин, — идет всегда избирательно, фрагментарно в отдельных органах, но всегда в крайней согласованности одного фрагмента с другим и всегда по принципу конечного создания работающей системы».[43] В частности, на примерах развития эмбрионов кошки, обезьяны и человеческого плода он показал функциональную обусловленность асинхронности развития отдельных структур зародышей, подчеркнув, что «в процессе эмбриогенеза идет ускоренное созревание отдельных нервных волокон, которые определяют жизненные функции новорожденного» (например, сосательные движения), ибо для его выживания «„система отношений“ должна быть полноценной к моменту рождения».[44]
Как показали исследования А. Д. Слонима и его сотрудников, внутриутробные движения влияют на координацию физиологических процессов, связанных с мышечной деятельностью, и тем самым способствуют подготовке поведения новорожденного. По данным Слонима, новорожденные козлята и ягнята способны, не утомляясь, бегать до двух часов подряд. Эта возможность обусловлена тем, что в ходе эмбриогенеза путем упражнения сформировалась координация всех функций, в том числе и вегетативных, необходимых для осуществления такой интенсивной деятельности уже в самом начале постнатального развития. В частности, к моменту рождения уже скоординирована регуляция минутного объема сердца и частота дыхания, как и других физиологических функций.
Говоря о принципе «конечного создания работающей системы» (Анохин) или эмбриональной преадаптации постнатального поведения, т. е. о преадаптационном значении эмбрионального поведения, нельзя забывать об относительности этих понятий. Эмбриональную преадаптацию нельзя понимать как некую изначальную, «фатальную» предопределенность. Условия жизни взрослого животного, его взаимодействие с компонентами окружающей среды в конечном итоге определяют условия развития зародыша, функционирование его развивающихся органов. Общие же закономерности и направление развития функции детерминированы сложившимися в филогенезе и генетически фиксированными факторами.
В указанной связи приобретает особый вопрос об «эмбриональном научении», считавшемся некоторыми исследователями преимущественным, если не единственным фактором всего сложного процесса первоначального формирования экзосоматических функций. К числу этих исследователей относится уже упоминавшийся известный американский ученый Цин Янг Куо, который еще в 20–30-х годах нашего века пытался объяснить весь процесс формирования поведения животных исключительно накоплением двигательного опыта и изменениями в окружающей зародыш среде. Куо сумел одним из первых убедительно показать, что уже в ходе эмбриогенеза происходит упражнение зачатков будущих органов, постепенное развитие и совершенствование двигательных функций путем накопления «эмбрионального опыта».
Куо изучал эмбриональное поведение на многих сотнях куриных зародышей (рис. 18). Чтобы получить возможность непосредственно наблюдать за движениями эмбрионов, он разработал и осуществил поистине виртуозные операции: перемещал зародыш внутри яйца, вставлял окошки в скорлупу и т. д. Ученый установил, что первые движения зародыша цыпленка, наблюдающиеся уже на четвертый, а иногда и на третий день инкубации, — это движения головы к груди и от нее. Уже через сутки голова начинает поворачиваться в стороны, и эти новые движения головы вытесняют прежние к 6–9-м суткам.
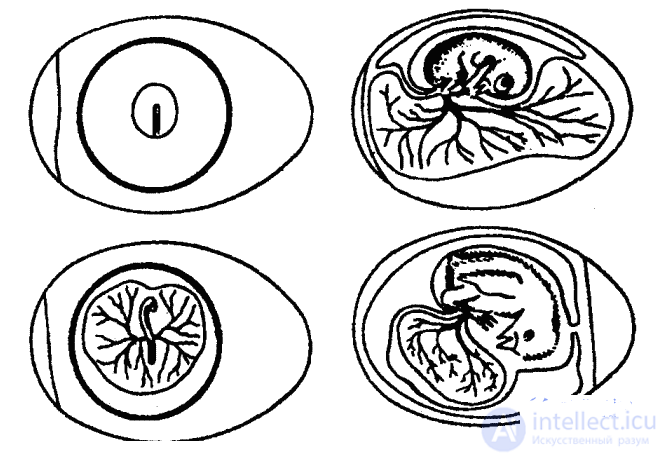
Рис. 18. Развитие куриного эмбриона. Сверху вниз: первые сутки инкубации (стадия первичной полоски — формируются зачатки головного и спинного мозга, скелета и скелетной мускулатуры), третьи сутки (зародыш на сосудистом поле желточного мешка), девятые сутки, пятнадцатые сутки
Очевидно, причина тому — отставание роста шейной мускулатуры от роста головы, которая к десятому дню составляет уже более 50 % веса всего зародыша. Такую огромную голову мышцы могут только поворачивать, но не поднимать и опускать. Кроме того, по Куо, движения головы формируются под влиянием таких моментов, как ее положение по отношению к скорлупе, расположение желточного мешка, сердцебиение и даже движения пальцев ног, поскольку последние во второй половине инкубационного периода располагаются слева и сзади головы.
Во всем этом Куо видел проявления действия «анатомического фактора» развития поведения. Сходным образом специфические условия морфоэмбриогенеза, анатомические изменения, связанные с ростом и развитием зародыша, влияют, по его мнению, и на формирование других движений куриного зародыша. В итоге вылупившийся из яйца цыпленок обладает целым набором выработанных в период эмбриогенеза реакций, «механизмов», но они не направлены на необходимые для поддержания жизни раздражители. Широкая генерализация характерна, по Куо, как для раздражителей (нет избирательного отношения к ним, на весьма различные раздражители следует одинаковая реакция), так и для двигательных реакций (всегда двигается все тело, движения отдельных его частей еще плохо или совсем не координированы, они нецелесообразны, неэкономичны).
Из своих исследований Куо вывел заключение, что цыпленок должен всему научиться, что ни одна его реакция не появляется в готовом виде, а следовательно, нет врожденного поведения. Сорок лет спустя после опубликования своих первых работ Куо, уточняя свою точку зрения, указал на то, что генетически фиксированные предпосылки формирования поведения могут по-разному реализоваться в зависимости от конкретных условий развития зародыша, но важнейшую роль играет в этом процессе «расшифровки» генетической информации отношение эмбриона к окружающей его среде. Одновременно Куо подчеркивает, что эмбриональное научение не следует рассматривать в традиционном аспекте, поскольку в развитии поведения эмбриона существенную роль играет самостимулирование. Однако, как показывают результаты современных исследований, тактильная и проприоцептивная стимуляция, как представлял себе ее Куо, играет в развитии моторики в нормальном эмбриогенезе, очевидно, подчиненную роль.
Односторонний подход к проблеме становления поведения в онтогенезе, игнорирование врожденной основы индивидуального поведения, в том числе на эмбриональной стадии развития, являются, конечно, глубоко ошибочными. Если и можно говорить о некоем научении в период эмбриогенеза, то оно не происходит на пустом месте, а является развитием и видоизменением определенного генетического зачатка, воплощением и реализацией в индивидуальной жизни особи видового опыта, накопленного в процессе эволюции. Филогенез подготовил возможность развития поведения в онтогенезе в биологически полезном для особи и вида направлении. Наследственность проявляется не только в строении организма, его систем и органов, но и в их функциях.
Наследственная основа эмбриогенеза поведения выступает особенно отчетливо в тех случаях, когда элементы поведения новорожденного проявляются сразу как бы в «готовом виде», хотя возможность предшествующего «эмбрионального научения» исключается. К таким случаям, очевидно, относятся у млекопитающих поиск соска и сосущие движения новорожденных, звуковые реакции и др. Здесь можно говорить лишь о пренатальном созревании функции без эмбрионального упражнения, т. е. без пренатальной функциональной тренировки соответствующих морфологических структур. Для такого созревания явно достаточно одной лишь врожденной программы развития, возникшей и закрепившейся в ходе эволюции вида.
Хорошим примером, показывающим наличие и роль генетически фиксированных «программ действия», может служить поведение новорожденного кенгуру, который появляется на свет на столь незрелой стадии развития, что его можно сравнить с эмбрионом высших млекопитающих. В известной степени можно считать, что окончательное развитие эмбриона завершается в сумке матери. Но несмотря на состояние крайнего незрелорождения, детеныш совершенно самостоятельно перебирается в сумку матери, проявляя при этом поразительные двигательные и ориентационные способности. При этом нахождение сумки осуществляется на основе отрицательного гидротаксиса (а возможно, и хемотаксиса): оказавшись вне родовых путей, новорожденный, цепляясь за шерсть, поднимается по сухим ее участкам к сумке, находит ее вход, вползает в нее, находит там сосок, крепко присасывается к нему и остается длительное время висеть на нем. Шерсть матери смачивается перед этим на прихвостовых участках туловища родовыми водами, выливающимися после разрыва зародышевых оболочек, «дорожка» же, ведущая к сумке, остается сухой.
Здесь обращает на себя внимание строгая последовательность врожденных реакций. На эмбриональном этапе своего развития, который здесь резко укорочен, если не сказать прерван, детеныш не мог научиться ни отдельным поведенческим актам этой цепи, ни этой последовательности. До разрыва родовых оболочек он постоянно находился во влажной среде и не мог, таким образом, упражнять отрицательное гидротаксисное поведение. Никогда он не соприкасался с сухими объектами, в том числе с шерстью. Аналогично обстоит дело и с другими компонентами этого сложного поведенческого комплекса, формирование которых также невозможно объяснить «пренатальным научением».
Существенным для понимания процессов созревания элементов поведения в эмбриогенезе является то обстоятельство, что у позвоночных иннервация соматической мускулатуры туловища и конечностей предшествует замыканию рефлекторных дуг (у куриного эмбриона это замыкание происходит уже на 6–7-й день с начала инкубации). Однако сокращения этих мышц начинаются уже с момента их иннервации и носят, таким образом, на первых порах нерефлекторный характер. Эти движения являются ритмичными, так как они обусловливаются спонтанными нейрогенными ритмами («импульсными ритмами»). Эндогенная ритмика в нервно-мышечных структурах сохраняется на протяжении всей жизни животного и представляет собой один из важных факторов созревания элементов врожденного поведения.
Все же едва ли можно говорить о некоем «чистом» созревании поведенческих актов, особенно если иметь в виду упомянутые выше коррелятивные морфофункциональные связи. К тому же, вероятно, никогда не удастся полностью исключить возможность и непосредственного пренатального упражнения тех или иных двигательных элементов поведенческого акта. Поэтому, возражая против одностороннего постулирования «эмбрионального научения» как единственного фактора пренатального развития поведения, невозможно соглашаться и с противоположной крайней точкой зрения, однобоко акцентирующей внимание на процессах эмбрионального (и постнатального) созревания. Эту точку зрения сформулировал видный американский исследователь онтогенеза поведения животных Л. Кармайкл, который допускал, что даже у человека поведение является на «девять десятых» врожденным. В том и другом случае неверным является противопоставление врожденного и приобретаемого в поведении, о чем уже многократно говорилось.
Что же касается термина «эмбриональное научение», то, очевидно, более точным будет выражение «эмбриональная тренировка», во всяком случае, когда речь идет о ранних стадиях эмбриогенеза. На первых стадиях, как уже упоминалось, даже еще нет рефлекторных дуг; сомнительно, чтобы условнорефлекторные связи образовались и на средних стадиях эмбриогенеза. Очевидно, отсутствуют на этих этапах и явления привыкания. Однако в каждом случае функционирование развивающегося органа или системы, безусловно, является индивидуальным «усвоением» и «прилаживанием» видового опыта, т. е. инстинктивного поведения, в виде тренировки. Последняя, разумеется, также относится к категории научения как одна из его элементарных форм. Полноценное же научение, как еще будет показано, встречается лишь на завершающих этапах эмбриогенеза.
Эмбриональное поведение беспозвоночных еще очень слабо изучено. Те немногие сведения, которые пока получены, относятся в основном к кольчатым червям, моллюскам и членистоногим. Известно, например, что зародыши головоногих моллюсков уже на ранних стадиях своего развития вращаются внутри яйца вокруг своей оси со скоростью один оборот в час. Об этом говорит сайт https://intellect.icu . В других случаях зародыши передвигаются от одного полюса яйца к другому. Все эти движения осуществляются с помощью ресничек, что представляет особый интерес, если учесть, что этот примитивный способ передвижения у взрослых головоногих моллюсков отсутствует, но широко распространен среди личинок морских беспозвоночных.
Заслуживает также внимания, что к концу эмбриогенеза у беспозвоночных оказываются уже вполне сформированными некоторые инстинктивные реакции, имеющие первостепенное значение для выживания. У мизид (морские ракообразные), например, к моменту вылупления уже вполне развита реакция избегания, т. е. уклонения от неблагоприятных воздействий. Но, как показал канадский исследователь М. Беррилл, первоначально у эмбриона наблюдается спонтанное «вздрагивание» и «подергивание», которые затем все чаще возникают в ответ на прикосновение к икринке. Таким образом, первоначально спонтанные движения постепенно сменяются рефлекторными.
У других ракообразных — морских козочек (отряд бокоплавы) — с 11-го по 14-й день развития, т. е. до вылупления, наблюдаются спонтанные и ритмичные движения головы и других частей эмбриона, из которых впоследствии формируются специфические двигательные реакции этих рачков. Только к концу эмбриогенеза, в день вылупления, появляются двигательные ответы на тактильные раздражения (прикосновения волоском к зародышу, с которого в эксперименте была снята яйцевая оболочка). В естественных условиях весь набор движений взрослой особи обнаруживается уже спустя 10 часов после вылупления.
У эмбриона дафнии антенны, служащие у взрослых особей для плавания, начинают двигаться на средних этапах эмбриогенеза, а перед его завершением они поднимаются и принимают то положение, которое необходимо для выполнения плавательных движений. В это время (за несколько часов до выхода из яйца) антенны начинают особенно интенсивно двигаться.
В этих примерах отчетливо проявляется постепенное формирование рефлекторного ответа на основе первоначально эндогенно обусловленных движений, которые впоследствии связываются с внешними раздражителями, отчасти уже путем «эмбрионального научения». Этот процесс сопряжен с глубокими морфологическими преобразованиями.
Первые движения зародышей рыб, по данным ряда исследователей, также возникают спонтанно на эндогенной основе. Еще в 20-х годах было показано, что движения зачатков органов появляются в строгой последовательности, в зависимости от созревания соответствующих нервных связей. После появления сенсорных нервных элементов на поведение зародыша начинают влиять и экзогенные факторы (например, прикосновения), которые сочетаются с генетически предопределенной координацией движений. Постепенно первоначально генерализованные движения зародыша дифференцируются.
Вообще у зародышей костистых рыб обнаруживаются к концу эмбриогенеза такие движения: дрожание, подергивание отдельных частей тела, вращение, змеевидное изгибание. Кроме того, перед вылуплением производятся своеобразные «клевательные» движения, облегчающие выход из яйцевидной оболочки. Кроме того, выклеву способствуют и изгибательные движения тела. В ряде случаев удалось установить четкую связь между появлением новых двигательных актов и общим анатомическим развитием.
Сходным образом совершается формирование эмбрионального поведения и у земноводных. Из первоначально генерализованного сгибания всего тела зародыша постепенно формируются плавательные движения, движения конечностей и т. д., причем и здесь двигательная активность развивается первично на эндогенной основе.
Интересный пример представляет в этом отношении жаба Eleutherodactylus martinicensis с острова Ямайка, у которой выход из икринки как бы задерживается и личинка развивается внутри яйцевых оболочек. Тем не менее у нее проявляются все движения, свойственные свободноплавающим личинкам (головастикам) других бесхвостых земноводных. Как и у последних, плавательные движения формируются у этой личинки постепенно из более генерализованных двигательных компонентов: первые движения конечностей еще слиты с общим извиванием всего тела, но уже спустя сутки можно вызвать одиночные рефлекторные движения одних конечностей независимо от движений мышц туловища; несколько позже и в строгой последовательности появляются более дифференцированные и согласованные движения всех четырех конечностей, и наконец возникают во всех деталях вполне координированные плавательные движения с участием всех соответствующих моторных компонентов, хотя плавать сформировавшаяся к этому времени личинка еще не начинала, ибо она по-прежнему заключена в яйцевые оболочки.
Что же касается хвостатых амфибий, то Когхилл показал в свое время, что эмбрион амбистомы производит плавательные движения еще задолго до вылупления, изгибаясь сперва наподобие буквы «С» и позже как буква «S». Еще позже появляются движения ног, типичные для передвижения по суше взрослой амбистомы, причем нейромышечная система, детерминирующая первичные плавательные движения, определяет и эти локомоторные элементы, особенно последовательность и ритм движений. Кармайкл сумел на этом же объекте доказать, что этот механизм созревает без научения. Он вырастил эмбриона амбистомы в анестезирующем растворе ацетонхлороформа, который полностью обездвиживал зародыш, но не препятствовал его росту и морфогенезу. В таких условиях локомоторные способности развивались вполне нормально, и в итоге они не отличались от таковых контрольных организмов, развитие которых совершалось в обычных условиях. Из своих опытов Кармайкл вывел заключение, что формирование способности к плаванию не нуждается в процессах научения, а зависит исключительно от анатомического развития.
Однако, как справедливо отмечал по поводу этих опытов польский зоопсихолог Я. Дембовский, у подопытных эмбрионов подавлялись лишь мышечные движения, возможность накопления двигательного эмбрионального опыта, но не другие функции, в частности процессы в развивающейся нервной системе. Невозможно, как пишет Дембовский, создать такие экспериментальные условия, при которых нервная система развивалась бы, не функционируя. Поскольку нервная система начинает функционировать еще до того, как она окончательно сформировалась, это функционирование также является своего рода упражнением, которое в свою очередь является важным фактором развития нервной деятельности, а тем самым всего поведения зародыша.
Для выявления эндогенной обусловленности формирования двигательной активности зародышей производились интересные опыты на эмбрионах саламандр: пересаживались зачатки конечностей таким образом, что последние оказывались повернутыми в обратную сторону. Если бы формирование их движений определялось эмбриональным упражнением (путем афферентной обратной связи), то в ходе эмбриогенеза должна была бы произойти соответствующая функциональная коррекция, восстанавливающая способности к нормальному поступательному движению. Однако этого не произошло, и после вылупления животные с повернутыми назад конечностями пятились от раздражителей, которые у нормальных особей обусловливают движение вперед. Сходные результаты были получены и у эмбрионов лягушек: перевертывание зачатков глазных яблок на 180° привело к тому, что оптокинетические реакции оказались у этих животных смещенными в обратном направлении (рис. 19).

Рис. 19. Инверсии оптокинетической реакции у лягушек (опыт Сперри). Большие черные стрелки обозначают направление оптического раздражения, вызывающего оптокинетическую реакцию (маленькая светлая стрелка) в норме (вверху) и после поворота зачатков глазных яблок на 1800 (внизу)
Все эти данные приводят к заключению, что формирование в эмбриогенезе локомоторных движений и оптомоторных реакций (а также некоторых других проявлений двигательной активности) происходит у низших позвоночных, очевидно, не под решающим влиянием экзогенных факторов, а в результате эндогенно обусловленного созревания внутренних функциональных структур.
Эмбриональное поведение птиц изучалось преимущественно на зародышах домашней курицы. Уже в конце вторых суток появляется сердцебиение, а начало спонтанной двигательной активности куриного эмбриона приурочено к 4-му дню инкубации. Весь период инкубации длится три недели. Движения начинаются с головного конца и постепенно простираются к заднему, охватывая все большие участки тела зародыша. Самостоятельные движения органов (конечностей, хвоста, головы, клюва, глазных яблок) появляются позже.
Как уже говорилось, Куо установил наличие и показал значение эмбриональной тренировки у зародыша курицы (а также других птиц), но впал при этом в односторонность, отрицая наличие врожденных компонентов поведения и спонтанной активности как таковой. В дополнение к сказанному приведем еще несколько примеров из работ этого выдающегося исследователя.
Куо установил, что максимальная двигательная активность эмбриона совпадает по времени с движениями амниона, т. е. внутренней зародышевой оболочки, обволакивающей зародыш. Из этого Куо заключил, что пульсация оболочки обусловливает начато двигательной активности последнего. Впоследствии, однако, В. Гамбургером было показано, что нет подлинной синхронизации между этими двигательными ритмами, а Р. В. Оппенгейм экспериментально доказал, что движения эмбриона не только не зависят от движения амниона, а, возможно, даже сами обусловливают их.
Формирование клевательных движений, по Куо, первично определяется ритмом биения сердца зародыша, ибо первые движения клюва, его открывание и закрывание, совершаются синхронно с сокращениями сердца. Впоследствии эти движения коррелируются со сгибательными движениями шеи, а незадолго до вылупления клевательный акт следует за любым раздражением тела в любой его части. Таким образом, реакция клевания, сформировавшаяся путем эмбриональной тренировки, имеет к моменту вылупления птенца весьма генерализованный характер. «Сужение» реакции в ответ на действие лишь биологически адекватных раздражителей происходит на первых этапах постэмбрионального развития. По Куо, так обстоит дело и с другими реакциями.
Важную роль в формировании эмбрионального поведения Куо отводил также непосредственному влиянию среды, окружающей зародыш, и происходящим в ней изменениям. Так, например, у куриного зародыша начиная с 11-го дня инкубации желточный мешок начинает надвигаться на брюшную сторону зародыша, все больше стесняя движения ног, которые до вылупления сохраняют согнутое положение, причем одна нога располагается поверх другой. По мере того как желток всасывается зародышем, верхняя нога получает все большую свободу движения, однако находящаяся под ней нога по-прежнему лишена такой возможности. Лишь после того как верхняя нога получит возможность достаточно далеко отодвигаться, нижняя может начать двигаться.
Таким образом, движения ног развиваются с самого начала не одновременно, а последовательно. Куо считал, что именно этим определяется развитие механизма последовательных движений ног цыпленка при ходьбе, что здесь кроется первопричина того, что вылупившийся из яйца цыпленок передвигается шагами, попеременно переставляя ноги, а не прыжками, отталкиваясь одновременно обеими лапками. Поддерживая концепцию Куо, Боровский писал по этому поводу, что «цыпленок шагает не потому, что у него имеется „инстинкт шагания“, а потому, что иначе не могут работать его ноги и их механизм, выросшие и развившиеся в таких именно условиях. Механизм этот не появится как нечто готовое в тот момент, когда он понадобится, а развивается определенным путем, под влиянием других факторов».[45]
Многие механистические концепции Куо не выдержали экспериментальной проверки, произведенной более поздними исследователями, другие еще нуждаются в такой проверке, тем более что вскрытые им коррелятивные отношения не обязательно указывают на причинные связи. Не подтвердилось, в частности, мнение Куо о том, что ведущим фактором двигательной активности зародыша в раннем эмбриогенезе является сердцебиение.
Гамбургером и его сотрудниками было установлено, что уже на ранних стадиях эмбриогенеза движения зародыша имеют нейрогенное происхождение. Электрофизиологические исследования показали, что уже первые движения обусловливаются спонтанными эндогенными процессами в нервных структурах куриного эмбриона. Спустя 3,5–4 дня после появления первых его движений наблюдались первые экстероцептивные рефлексы, однако, Гамбургер, Оппенгейм и другие показали, что тактильная, точнее, тактильнопроприоцептивная стимуляция не оказывает существенного влияния на частоту и периодичность движений, производимых куриным эмбрионом на протяжении первых 2–2,5 недель инкубации. По Гамбургеру, двигательная активность зародыша на начальных этапах эмбриогенеза «самогенерируется» в центральной нервной системе.
Гамбургером производился следующий опыт: перерезав зачаток спинного мозга в первый же день развития куриного эмбриона, он регистрировал впоследствии (на 7-й день эмбриогенеза) ритмичные движения зачатков передних и задних конечностей. Нормально эти движения протекают синхронно. У оперированных же эмбрионов эта согласованность нарушилась, но сохранилась самостоятельная ритмичность движений.
Это указывает на независимое эндогенное происхождение этих движений, а тем самым и соответствующих нервных импульсов, на автономную активность процессов в отдельных участках спинного мозга. С развитием головного мозга он начинает контролировать эти ритмы. Вместе с тем эти данные свидетельствуют о том, что двигательная активность не обусловливается исключительно обменом веществ, например, такими факторами, как уровни накопления продуктов обмена веществ или снабжения тканей кислородом, как это принималось некоторыми учеными.
При изучении эмбрионального развития поведения птиц необходимо учитывать специфические особенности биологии исследуемого вида, которые отражаются и на протекании эмбриогенеза. Особенно это касается различий между выводковыми (зрелорождающимися) и птенцовыми (незрелорождающимися) птицами. Так, например, как показал советский исследователь Д. Н. Гофман, по сравнению с курицей грач развивается более ускоренно, быстрее накапливается масса тела зародыша, зато у курицы эмбриогенез проходит более равномерно и имеется больше периодов роста и дифференциации. Последний период формирования морфологических структур и поведения проходит у курицы еще внутри яйца, у грача же (как не зрелорождающейся птицы) этот период относится к постэмбриональному развитию.
В отличие от рассмотренных до сих пор животных зародыши млекопитающих развиваются в утробе матери, что существенно усложняет (и без того весьма трудное) изучение их поведения, поэтому по эмбриональному поведению млекопитающих накоплено значительно меньше данных, чем по куриному эмбриону и зародышам земноводных и рыб. Непосредственные визуальные наблюдения возможны лишь на эмбрионах, извлеченных из материнского организма, что резко искажает нормальные условия их жизни. Рентгенологические исследования указывают на то, что двигательная активность таких искусственно изолированных зародышей выше, чем в норме. Вместе с тем именно на таких объектах, преимущественно зародышах грызунов, были получены те данные, которыми мы сегодня располагаем.
Так, например, по Кармайклу, развитие двигательной активности совершается у зародыша морской свинки следующим образом. Первые движения состоят в подергивании шейно-плечевого участка туловища эмбриона. Они появляются приблизительно на 28-й день после оплодотворения. Постепенно появляются и другие весьма разнообразные движения, а к 53-му дню, т. е. приблизительно за неделю до родов, формируются четко выраженные реакции, которые достигают максимального развития за несколько дней до рождения. У такого эмбриона обнаруживаются уже вполне адекватные, и главное, видоизменяющиеся рефлекторные ответы на тактильные раздражения: прикосновение волоском к коже около уха вызывает специфическое подергивание последнего, непрерывное продолжение этого раздражения или его многократное повторение — приведение конечности данной стороны к раздражителю, чем нередко достигается его удаление; если же и после этого продолжать раздражение, то приходит в движение вся голова, а затем и туловище эмбриона, что может привести к его вращению, и наконец, по выражению Кармайкла, «каждая мышца приводится в действие». Кармайкл говорил по этому поводу, что эмбрион ведет себя как бы согласно пословице: «если не сразу достиг успеха — пробуй, пробуй еще», но подчеркивал, что нет оснований предполагать, чтобы что-либо в этом поведении являлось выученным.
Эмбриональное развитие поведения млекопитающих существенно отличается от такового у других животных. Это отличие выражается в том, что у млекопитающих движения конечностей формируются не из первоначальных общих движений всего зародыша, как мы это видели у вышеупомянутых других позвоночных, особенно низших, а появляются одновременно с этими движениями или даже раньше их. Вероятно, в эмбриогенезе млекопитающих большее значение приобрела ранняя афферентация, чем спонтанная эндогенная нейростимуляция.
Постоянная тесная связь развивающегося зародыша с материнским организмом, в частности посредством специального органа — плаценты, создает у млекопитающих совершенно особые условия для развития эмбрионального поведения. Новым и весьма важным фактором является в этом отношении возможность воздействия на этот процесс со стороны материнского организма, прежде всего гуморальным путем.
На такую возможность косвенно указывают результаты экспериментов, при которых на женских зародышей морской свинки еще во время их внутриутробного развития воздействовали мужским половым гормоном (тестостероном). В результате, став половозрелыми, они проявили признаки самцового поведения в ущерб сексуальному поведению, свойственному нормальным самкам. Аналогичное воздействие, произведенное после рождения, не давало такого эффекта. Подобным образом удавалось преобразовывать и половое поведение мужских особей. Очевидно, в ходе эмбриогенеза содержание тестостерона в организме зародыша влияет на формирование центральных нервных структур, регулирующих сексуальное поведение: его отсутствие — в сторону женских признаков, его наличие — в сторону мужских.
В экспериментах ряда исследований у беременных самок крыс периодически вызывали состояния беспокойства. В таких условиях родились более пугливые и возбудимые детеныши, чем в норме, несмотря на то, что их затем вскармливали другие самки, не подвергавшиеся экспериментальным воздействиям. Эти данные особенно отчетливо показывают роль влияния материнского организма на формирование признаков поведения детеныша в эмбриональном периоде его развития.
Выше приводились примеры рефлекторных движений зародышей, производимых преимущественно в ответ на тактильные раздражения. Сенсомоторная активность составляет единый процесс на всех этапах жизни животного, хотя, как мы видели, двигательный компонент является в эмбриогенезе первичным и может возникнуть на эндогенной основе. Вместе с тем, по мере развития эмбриона и формирования его рецепторных систем, все большее значение приобретает сенсорная стимуляция, выступающая, очевидно, также в форме самостимуляции.
Куо видел такую самостимуляцию, в частности, в том, что куриный зародыш прикасается одной частью тела (например, ногой или крылом) к другой части (например, голове) и вызывает тем самым двигательную реакцию последней. Оппенгейм, правда ссылаясь на собственные исследования и работы других авторов, ставит под сомнение правомерность выводов Куо о таком механизме самостимуляции, но не отрицает существования эмбриональных сенсомоторных связей, как и значения сенсорной стимуляции в эмбриональном поведении.
Еще в начале 30-х годов Д. В. Орр и В. Ф. Уиндл сумели показать, что наряду со спонтанной двигательной активностью у куриного эмбриона развивается рефлекторная система движений. Изолированные движения крыла возникают в ответ на тактильное раздражение уже на ранних стадиях эмбриогенеза; это указывает на то, что потенциальные возможности рефлекторных реакций существуют уже тогда, когда еще отсутствуют реальные возможности внешней аффектации и двигательная активность зародыша проявляется лишь в общих спонтанных телодвижениях. Эти же ученые установили, что у куриного эмбриона моторные структуры нервной системы формируются раньше сенсорных, а первые реакции на внешние раздражения появляются лишь через четыре дня после первых спонтанных движений.
Однако наибольшее значение сенсорная стимуляция приобретает у куриного зародыша на последних стадиях эмбриогенеза, за 3–4 дня до вылупления (Гамбургер). Именно в этот период в развитие поведения включаются у птиц как мощные внешние факторы оптические и акустические стимулы, подготавливающие птенцов к биологически адекватному общению с родительскими особями.
Зрение и слух появляются лишь к концу эмбриогенеза и не влияют на развитие ранней двигательной активности зародыша. Правда, как было установлено рядом советских исследователей (Т. П. Блинкова, Г. Е. Свидерская и др.), сильные внешние раздражения способны вызвать реакции куриного зародыша уже на средних и даже ранних стадиях эмбриогенеза. Реакции на громкие звуки обнаруживаются не только после 14–19-го дня, когда уже начинает функционировать орган слуха, но и даже начиная с 5-го дня инкубации. В это же время можно вызвать реакции и на мощные световые воздействия. Все эти реакции выражаются в усилении или торможении эмбриональных движений. Однако, не говоря уже о том, что в данных экспериментах зародыши подвергались экстремальным, биологически неадекватным воздействиям, свет и звук могут на этом этапе выступать лишь как физические агенты, непосредственно влияющие на мышечную ткань или кожу, но не как носители оптической или акустической информации.
Если же, как явствует из новых данных американского ученого Г. Готтлиба, воздействовать на зародыш биологически адекватными, т. е. обычно встречающимися в природе, звуками на такой стадии, когда он еще не реагирует на подобные раздражения, то это может положительно сказаться на появляющихся позже слуховых реакциях эмбриона.
Что касается развития оптических реакций, то только начиная с 17–18-х суток инкубации в глазу и зрительных долях куриного эмбриона обнаруживаются электрофизиологические изменения в ответ на оптические раздражения. У зародыша пекинской утки, например, зрачковый рефлекс появляется на 16-й день инкубации, но это чисто фотохимическая реакция, которая не имеет функционального значения и сменяется на 18-й день (т. е. относительно раньше, чем у куриного эмбриона) подлинно нервной реакцией. Очевидно, к этому времени уже функционируют периферические и центрально-нервные элементы зрительного анализатора. Перед вылуплением птенцов зрачковый рефлекс практически уже так же развит, как у взрослой утки.
У эмбрионов многих птиц в последние дни перед вылуплением не только начинают вполне функционировать дистантные рецепторы, т. е. органы зрения и слуха, но и появляются первые активные действия, направленные на внешнюю среду, а именно подача сигналов насиживающим родительским особям. Так, например, у представителя отряда чистиков кайры птенец научается еще за 3–4 дня до вылупления отличать голос родительской особи от голосов других кайр, гнездящихся в тесном соседстве на птичьих базарах. Если передискусственно инкубируемыми яйцами проигрывать магнитофонную запись криков какой-либо определенной взрослой кайры, а затем воспроизводить эту запись одновременно с записью криков другой кайры перед вылупившимися из этих яиц птенцами, то они направятся в сторону звуков, которые слышали еще до вылупления. Контрольные же птенцы из «неозвученных» яиц направятся в промежуток между источниками звуков, а затем начнут метаться между ними. Было установлено, что распознавание родительского голоса (в отличие от голосов соседних птиц) осуществляется на основе согласования ритмов подачи звуков родителя и невылупившегося птенца: в ответ на писк последнего насиживающая птица приподнимается, передвигает яйцо и сама подает голос. Таким образом, кинестетические ощущения эмбриона сочетаются с акустическими, а в целом его активность совпадает с таковой взрослой птицы, что и позволяет установить принадлежность услышанного звука родительской особи (исследования Б. Чанц).
В этом примере мы видим, как созревшее в эмбриогенезе врожденное, инстинктивное поведение (локомоторная реакция вылупившихся в изоляции птенцов на видоспецифический звук, т. е. на ключевой раздражитель) сочетается с истинным эмбриональным научением (условнорефлекторным путем), результатом которого является индивидуальное опознавание еще в пренатальном периоде развития, дифференцировка индивидуальных отличий видотипичных звуков. Именно в этом направлении совершается достройка соответствующего врожденного пускового механизма птенца, его обогащение необходимыми дополнительными признаками путем научения (в данном случае пренатального). Мы еще встретимся с этим вопросом при рассмотрении процессов запечатления. Сейчас же нам важно отметить, что между эмбрионом и родительской особью происходит обмен сигналами и возникает первичная коммуникация.
Сходные результаты были получены и у других видов птиц, в том числе у близкой родственницы кайры гагарки. Пренатальное распознавание голосов родителей было установлено также у гусиных (канадская казарка), куликов (кулик-сорока) и представителей других отрядов птиц. В частности, сравнение поведения птенцов, выведенных в инкубаторе в условиях звуковой изоляции, с таковым птенцов, слышавших крики взрослых птиц своего вида незадолго до вылупления, показало существенное значение эмбрионального научения и у чаек. Немецкая исследовательница М. Импековен установила, что изменения в поведении взрослых чаек (Larus atricilla) при переходе от насиживания к уходу за птенцами в большой степени обусловливаются акустическими сигналами, подаваемыми птенцами перед выклевом. При таких звуках насиживающая птица начинает посматривать вниз, на яйца, попеременно вставать и садиться, перекатывать яйца, отряхиваться и издавать ответные крики. Эти реакции удалось вызвать, воспроизводя издаваемые птенцом звуки в «чистом виде», т. е. в магнитофонной записи. При их замедленном проигрывании, т. е. изменении физических параметров звуков, реакции родительской особи ослабевают и появляются реже. С другой стороны, Импековен удалось экспериментально доказать, что родительские крики, услышанные птенцами еще до вылупления, стимулируют клевательные движения последних, в том числе и постнатальное клевание клюва родительской особи, т. е. «попрошайничество». Таким образом, стимулирующее влияние родительских криков проявляется здесь как результат пренатального накопления опыта.
Как видно из изложенного, в эмбриогенезе происходит интенсивная подготовка к последующим, постнатальным этапам формирования поведения, а отчасти и само формирование элементов поведения новорожденного путем, с одной стороны, развития генетически обусловленных компонентов активности и, с другой стороны, накопления эмбрионального опыта. Как и в постнатальной жизни животного, эти две стороны единого процесса развития поведения — врожденное и приобретаемое — невозможно отрывать друг от друга и изучать вне их взаимосвязи, поэтому неверно рассматривать эмбриогенез поведения с точки зрения альтернативы: созревание врожденных элементов поведения или эмбриональное упражнение. В каждом случае речь может идти лишь о том, какой из этих компонентов превалирует или даже доминирует. И здесь сложным образом переплетаются воздействия различных факторов в разных соотношениях и сочетаниях, как это происходит и на постнатальном этапе развития.
При этом необходимо, естественно, учитывать те специфические особенности, которыми отличается пренатальное развитие поведения животных. Это касается прежде всего роли внешней среды в формировании пренатальной двигательной активности и психической деятельности.
Приведенные выше данные показывают, что развитие поведения в пренатальном периоде онтогенеза совершается у низших и высших животных неодинаково, хотя и обнаруживает ряд общих черт. Эти филогенетические различия обусловлены закономерностями эволюции эмбриогенеза, установленными прежде всего Северцовым, о чем уже говорилось выше. Но в целом можно сказать, что у всех животных — во всяком случае на ранних стадиях эмбриогенеза — прямые влияния внешней среды играют незначительную роль (или не играют вообще никакой роли) в формировании отдельных форм двигательной активности.
Средой, в которой развивается эмбрион млекопитающего, является материнский организм, который не только хранит и защищает его от неблагоприятных воздействий, но и непосредственно обеспечивает всю его жизнедеятельность. Поэтому утроба является средой обитания зародыша, на которую направлена его активность. Однако связь эмбриона с подлинным внешним миром, в котором протекает вся постнатальная жизнь развивающегося организма, осуществляется лишь опосредованно через организм матери и не может иметь существенного значения для развития психики млекопитающего во внутриутробном периоде его развития.
В отличие от млекопитающих и, вероятно, других живородящих животных при внеутробном развитии зародыш часто подвергается воздействиям различных агентов внешней среды. Однако, как уже отмечалось, экспериментальные исследования показали, что эти агенты едва ли могут непосредственно направлять развитие первичных форм двигательной активности эмбриона. Но если даже допустить обратное, а именно что такие воздействия в состоянии оказать прямое влияние на формирование этой активности, то такая связь по необходимости была бы односторонней, ибо сфера двигательной активности зародыша не простирается дальше яйцевых оболочек и не может вызвать физические изменения в окружающей среде. Но это означает, что отсутствует важнейший источник психической деятельности.
Другое обстоятельство, крайне лимитирующее возможности развития психики в эмбриональном периоде развития, — это однородность, постоянство и бедность компонентами среды, которая окружает зародыша как в яйце (птичьем или икринке), так и в утробе млекопитающего. Там ему практически «нечего отражать». Поэтому будет, очевидно, правильно сказать, что психика эмбриона — это психика в процессе ее становления. Эмбрион — это еще не полноценное животное, а формирующийся организм животного на начальном этапе своего развития. Животная жизнь невозможна без активного взаимодействия с внешней (т. е. постнатальной) средой, а как раз это взаимодействие еще отсутствует на эмбриональном этапе развития, по меньшей мере на его ранних стадиях. В ходе эмбриогенеза осуществляется лишь подготовка к этому взаимодействию.
На ранних стадиях эмбриогенеза формируются предпосылки, потенциальные возможности психического отражения, т. е. существуют только зачаточные формы элементов психики. Лишь по мере того как формируются органы и системы органов развивающегося организма и появляется необходимость установления и расширения связей с внешним миром, зарождается и развивается психическое отражение, которое является функцией этих структур и служит установлению этих связей.
Как было показано, это происходит в конце эмбриогенеза, во всяком случае у птиц, у которых наряду с весьма дифференцированной двигательной активностью (и на ее основе) появляется коммуникативное поведение, обеспечивающее установление контактов с внешним миром (конкретно — с родительской особью) еще до вылупления.
Во взаимосвязях между невылупившимся птенцом и родительской особью, в согласовании их поведения на протяжении последних дней инкубации проявляется уже достаточно сложная психическая деятельность эмбриона. Правда, это относится лишь к заключительной стадии пренатального онтогенеза. При этом необходимо также учесть, что такая психическая активность зародыша едва ли свойственна млекопитающим, у которых эмбриональное развитие является по сравнению с птицами относительно укороченным: детеныши рождаются на более ранних стадиях эмбриогенеза, на которых можно предположить наличие лишь примитивных элементов будущей психической активности.
Итак, можно сказать, что значение эмбриогенеза для формирования психической деятельности состоит в том, чтобы подготовить морфофункциональную основу психического отражения. Это относится как к двигательным компонентам психической деятельности, так и к подготовке условий для функционирования сенсонейромоторных систем на постэмбриональном этапе развития.
Ясно, что будет поздно, если эти предпосылки начнут формироваться лишь после появления животного на свет, поэтому такая база должна уже существовать к началу постнатального развития, чтобы организм мог приступить к построению всесторонних отношений с компонентами среды его обитания. Для подготовки этой базы достаточна генетически фиксированная спонтанная двигательная активность эмбриона, дополняемая более или менее выраженным эмбриональным научением. Такая активность может формироваться и в той константной, однообразной, нерасчлененной, бедной предметными компонентами среде, в которой живет и развивается зародыш.
Если я не полностью рассказал про развитие психической деятельности в пренатальном периоде? Напиши в комментариях Надеюсь, что теперь ты понял что такое развитие психической деятельности в пренатальном периоде и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Сравнительная психология и Зоопсихология
Комментарии
Оставить комментарий
Сравнительная психология и Зоопсихология
Термины: Сравнительная психология и Зоопсихология