Лекция
Привет, мой друг, тебе интересно узнать все про перцептивная психика, тогда с вдохновением прочти до конца. Для того чтобы лучше понимать что такое перцептивная психика , настоятельно рекомендую прочитать все из категории Сравнительная психология и Зоопсихология.
Низший уровень развития перцептивной психики
перцептивная психика является высшей стадией развития психического отражения. Напомним, что эта стадия характеризуется, по Леонтьеву, изменением строения деятельности — выделением содержания деятельности, относящейся к условиям, в которых дан объект деятельности в среде (операции). По этой причине мы встречаемся здесь уже с подлинными навыками и восприятиями. Предметные компоненты среды отражаются уже как целостные единицы, в то время как при элементарной сенсорной психике имело место отражение лишь отдельных их свойств или суммы последних. Предметное восприятие обязательно предполагает определенную степень обобщения, появляются чувственные представления.
Разумеется, перцептивная психика, свойственная огромному числу животных, стоящих на разных ступенях эволюционного развития, обнаруживает в конкретных своих проявлениях большие различия. Поэтому необходимо и на этой стадии развития психики выделить отдельные уровни, из которых здесь будет рассмотрен низший.
На этом уровне находятся прежде всего высшие беспозвоночные — головоногие моллюски и членистоногие (по меньшей мере высшие). Характеристику рассматриваемого уровня психического развития мы дадим на примере насекомых — центрального класса типа членистоногих. (Кроме насекомых к членистоногим относятся ракообразные, паукообразные, многоножки и некоторые мелкие группы.) Насекомые — самый многочисленный класс животных как по количеству видов, так и по количеству особей. Насекомые обитают повсюду на суше (во всех климатических зонах) — как на поверхности, так и в почве, во всех пресных водоемах, а также в воздухе, поднимаясь на высоту до двух километров.
Движения
Огромное разнообразие «экологических ниш», занимаемых насекомыми и другими членистоногими, обусловило, естественно, развитие весьма различных форм приспособления в области строения и поведения этих животных. Наиболее существенным и характерным для эволюции членистоногих, особенно насекомых, было завоевание новой среды обитания — суши. Членистоногие — первые наземные животные в истории Земли. Переход на сушу был сопряжен с развитием особых органов передвижения — конечностей в виде сложных рычагов, состоящих из отдельных, соединенных суставами члеников, по которым членистоногие и получили свое название. Такие сложные рычаги (их аналогами являются конечности наземных позвоночных) наилучшим образом служат для передвижения по твердому субстрату путем отталкивания от него. Членистоногие в состоянии передвигаться по суше почти всеми возможными способами (ползание, ходьба, бег как по горизонтальным, так и по вертикальным поверхностям, прыгание, передвижение в толще грунта). В воде членистоногие плавают, ныряют, ползают и даже бегают по ее поверхности.
У насекомых, кроме того, в связи с переходом к жизни в воздушной среде впервые появился и совершенно новый способ передвижения — полет с помощью крыльев. Насекомые способны развить в полете очень большую скорость: у некоторых стрекоз она достигает 90 километров в час, у самца овода — даже 122 километра в час. Поистине поразительна частота взмахов крыльев у многих насекомых. Так, мотылек делает лишь 9 ударов в секунду, пчела же — 190, а комнатная муха — 330, но у некоторых мух и перепончатокрылых скорость вибрации крыльев достигает до тысячи ударов в секунду. Столь быстрые движения и соответственно частоты мышечных сокращений не наблюдаются ни у каких других животных.
Соответственно и крылевая мускулатура насекомых отличается исключительно сложным устройством: она состоит из множества различно располагающихся мышечных пучков, частично действующих как антагонисты. Большую сложность обнаруживают и другие отделы мышечной системы.
Вообще у насекомых (не считая крылевой мускулатуры насекомых) различают следующие отделы мышечной системы: мышцы туловища, заведующие его движениями: мышцы конечностей, с помощью которых движется вся конечность по отношению к туловищу; мышцы отдельных члеников конечностей, мышцы внутренних органов.
Как видно, мускулатура насекомых отличается высокой степенью дифференцированности и специализированности. К тому же подавляющее большинство их мышц — поперечнополосатые. Количество отдельных мускулов исключительно велико — полторы тысячи и более. Внушительна и сила мышц: насекомые способны таскать предметы, иногда в сто раз превышающие их собственный вес. В целом, как отмечал Догель, у членистоногих мышечная система беспозвоночных достигает «апогея своего развития» как в морфологическом, так и в физиологическом отношении.
Центральная нервная система
В соответствии со сложной и высокодифференцированной организацией двигательного аппарата находится и сложное строение центральной нервной системы насекомых, которую, однако, мы можем здесь охарактеризовать лишь в самых общих чертах.
Как и у кольчатых червей, основу всей нервной системы составляют у насекомых метамерно расположенные ганглии, связанные друг с другом межсегментными нервными стволами. Эти стволы сближаются у большинства насекомых настолько, что образуют (хотя бы внешне) одну непарную нервную цепочку. Одновременно имеет место и слияние последовательных метамерных ганглиев.
В связи с отчетливо выраженной цефализацией в головном отделе слившиеся ганглии образовали мощный головной мозг в виде сложно устроенного надглоточного узла, в котором различают три отдела: протоцеребрум, дейтоцеребрум и тритоцеребрум. Передний отдел, протоцеребрум, развит сильнее остальных. Особое внимание заслуживают находящиеся в нем так называемые грибовидные (или стебельчатые) тела — ассоциативные и координирующие центры. В наибольшей степени они развиты у насекомых с особенно сложным поведением — у пчел, прежде всего рабочих особей, и муравьев. С протоцеребрумом связана и пара очень крупных и сложно устроенных зрительных долей, иннервирующих сложные глаза. Дейтоцеребрум (средний отдел головного мозга) иннервирует усики, тритоцеребрум иннервирует область рта и, кроме того, тесно связан с симпатической нервной системой. Вообще для насекомых характерна большая концентрация нервных элементов. Такая концентрация повышает возможности управления отдельными системами органов и поднимает общий морфофизиологический уровень организма как целостной единицы.
Нервная система, и в частности головной мозг, устроена у разных членистоногих, однако, весьма различно. Существенные различия обнаруживаются между низшими и высшими представителями даже в пределах классов, а иногда и отрядов. Так, например, отношение размеров мозга к объему туловища равняется у майского жука 1: 3290, у муравья 1: 280, а у пчелы 1: 174.
На очень высоком уровне развития находится нервная система головоногих моллюсков, в корне отличаясь от таковой других моллюсков. Особенно это относится к их весьма сложно устроенному головному мозгу. Обращает на себя внимание исключительная концентрация мозговых ганглиев: ганглии слились воедино и образовали общую околопищеводную массу. Основания зрительных нервов образуют громадные вздутия — оптические ганглии, которые своими размерами нередко превосходят сами церебральные узлы.
Нервы, отходящие от мозга к эффекторам, достигают очень большой мощности — до миллиметра и больше в диаметре, что в 5–10 раз превышает толщину нервов большинства позвоночных! Этим обеспечивается значительно большая скорость проведения нервных импульсов, чем у низших моллюсков. Соответственно огромной величине глаза исключительно развит и зрительный нерв. Выдающийся советский гистолог А. А. Заварзин показал большое сходство тонкого гистологического строения оптических центров головоногих и позвоночных.
Таким образом, мы видим, что глубокие различия между элементарной сенсорной и перцептивной психикой находят свое воплощение и в структуре нервной системы уже на низшем уровне перцептивной психики.
Сенсорные способности, таксисы
Специфические условия, в которых живут насекомые, далеко идущее приспособление двигательной активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением, — все это обусловило появление у данных животных многих, и притом своеобразно устроенных, органов чувств.
Большую роль в жизни насекомых играет зрение. В разном количестве у разных видов имеются простые одиночные глаза. Но главный орган зрения — своеобразный сложный глаз, получивший название фасеточного, так как его поверхность состоит из отдельных, четко ограниченных участков — фасеток. Каждой фасетке соответствует самостоятельный глазок — омматидий, изолированный от своих соседей пигментной прокладкой и функционирующий независимо от них. Общий внешний покров глаза образует над омматидиями утолщения, обозначаемые как роговица или роговой хрусталик и играющие роль диоптрического аппарата глаза. Каждый омматидий содержит 7–8 зрительных клеток, аксоны которых оканчиваются в оптических ганглиях мозга.
Благодаря такому устройству сложных глаз в мозг насекомого передаются «мозаичные» изображения воспринимаемых предметных компонентов среды. Конечно, это не означает, что насекомое действительно видит окружающий мир в виде мозаики. В мозгу поступающие из отдельных омматидиев зрительные раздражения синтезируются в целостную картину, как это имеет место и у позвоночных. Все же нам очень трудно представить себе, как на самом деле видит насекомое.
Еще в прошлом веке делались попытки установить путем фотографирования через фасеточные глаза четкость и дальность зрения насекомых и раков. Позже, в частности в 60-х годах нашего века, такие опыты повторялись, но едва ли возможно таким образом решить проблему, ибо, как известно, видит не глаз, а мозг. Насекомые, очевидно, весьма близоруки: ясное видение, например, бабочки-крапивницы возможно лишь на расстоянии приблизительно 2–3 сантиметров. От объектов, расположенных дальше (уже начиная с 5–7 сантиметров), насекомые получают лишь расплывчатые изображения, но, очевидно, этого достаточно для их общей ориентировки в пространстве.
На расстоянии пчела способна увидеть лишь объекты в сто раз большей величины, чем это необходимо для зрения человека. Отдельный цветок она увидит лишь тогда, когда она приблизится к нему вплотную. Все эти особенности зрения насекомых вполне соответствуют тому микроландшафту, в котором живут эти животные.
Острота зрения (разрешающая способность, т. е. способность видеть две сближенные точки) зависит у насекомых при прочих равных условиях, в частности, от количества омматидиев, которое, однако, сильно колеблется даже в пределах отрядов. Так, в отряде жуков это количество колеблется от трех-четырех десятков до нескольких десятков тысяч. У пчелы каждый глаз состоит приблизительно из четырех тысяч фасеток. Чем мельче фасетки, тем слабее в них освещение, но тем детальнее зрение. Особенно большое значение это имеет для предметного зрения. Кроме того, при более выпуклом глазе оси омматидиев сильнее расходятся, и в результате расширяется поле зрения. Одновременно суживается точка, видимая одной фасеткой, чем также увеличивается острота зрения.
Наилучшим образом насекомые видят подвижные объекты или неподвижные предметы во время собственного движения. Пролетающая над лугом пчела, например, видит покачивающиеся на ветру цветы или же изображение неподвижного цветка скользит по ее фасеточному глазу. В любом случае это подвижное (расплывчатое) изображение заставляет ее спуститься вниз и обследовать растение с минимального, т. е. оптимального для зрения пчелы, расстояния.
Что касается цветового зрения, то видимый ими спектр шире, чем у человека, так как его граница проходит в ультрафиолетовом секторе. Спорный вопрос о способности насекомых к восприятию красного цвета в настоящее время, очевидно, решен в положительном смысле (за исключением медоносной пчелы и некоторых других насекомых). Советский энтомолог Г. А. Мазохин-Поршняков считает, что вообще цветовое зрение у взрослых насекомых представляет универсальное явление. По его данным, цвета различаются даже видами, активная жизнь которых протекает ночью.
До сих пор мы касались только реакции насекомых на свет, который играет в их жизни очень большую роль. Не меньшее, если не большее значение имеют для них запахи. В естественном поведении насекомых стимулы различной модальности всегда выступают в комплексе. Так, уже упомянутые осы рода Philanthus охотятся за пчелами, руководствуясь зрением с расстояния 30 сантиметров, но опознают их по запаху лишь с расстояния 15–5 сантиметров. Во время зрительного поиска оса совершенно не реагирует на запах пчелы.
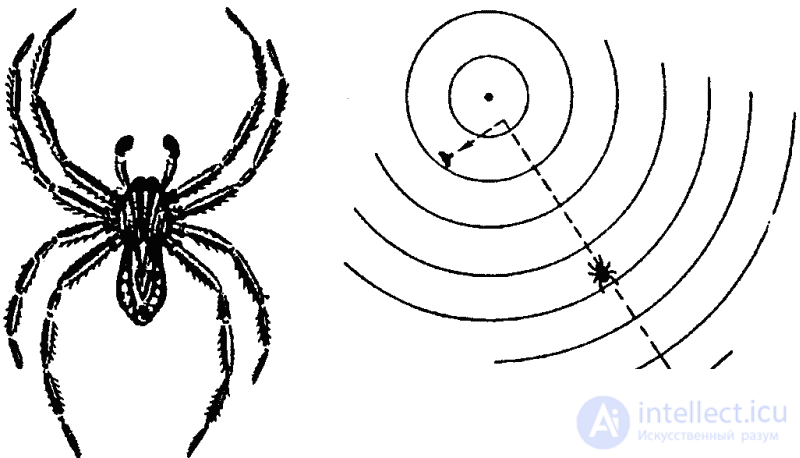
Аналогично обстоит дело и у других членистоногих. Водяные пауки рода Pirata ориентируются прежде всего по вибрации поверхности воды, в непосредственной же близости от жертвы — зрением (рис. 36). Другие пауки узнают о попадании в их сеть насекомого, воспринимая вибрации паутины. Но обследование жертвы производится сразу тремя способами: паук прикасается к ней передними конечностями, снабженными хеморецепторами, а также так называемыми щупиками (органы осязания), и, кроме того, паук на вкус пробует жертву укусом.
Для ознакомления с таксисами у насекомых рассмотрим в качестве примера фототаксисы. Эти таксисы (положительные или отрицательные) встречаются у насекомых прежде всего в виде тропо-, тело- и менотаксисов, хотя не утратили своего значения и примитивные таксисы. Так, например, положительные и отрицательные клинотаксисы с четко выраженными маятникообразными поисковыми движениями играют ведущую роль в оптической ориентации у многих личинок мух. Тропо- и телотаксисы связаны у них друг с другом многими переходными формами. У взрослых насекомых чаще всего встречаются, однако, четко выраженные телотаксисы. Как правило, тропотаксисное движение очень скоро переходит у них в телотаксисное. У комаров и вшей преобладает тропотаксисная ориентация движения.
Менотаксисы, относящиеся к более сложным формам ориентации, встречаются у высших насекомых (муравьев, пчел, бабочек, жуков). Немецкий исследователь поведения насекомых М. Линдауер выделяет здесь следующие четыре случая: 1) угол фиксации источника света случаен (муравьи, жук-навозник); 2) угол фиксации является врожденным (некоторые бабочки); 3) угол фиксации сообщается особями одного вида друг другу («танцы» пчел, информирующие об угле ориентации по положению солнца); 4) угол фиксации выучивается в индивидуальном порядке. Последний случай можно уже отнести к высшей категории таксисов — мнемотаксисам (выученная ориентация), которые широко распространены среди позвоночных, но также встречаются у высших насекомых (пчел, ос, муравьев) (рис. 37).

Необходимо, однако, подчеркнуть, что и в тех случаях, когда ориентация осуществляется на основе других фототаксисов, поведение насекомых одновременно определяется рядом дополнительных факторов. Например, доказано, что у многих насекомых характер их фототаксисного поведения сильно видоизменяется в зависимости от температуры воздуха, времени дня, физиологического состояния и т. д. Хорошо известно, что муха не реагирует на свет, пока спокойно летает по комнате. Но стоит начать ее преследовать, как у нее сейчас же «просыпается» положительный фототелотаксис: она быстро направляется к источнику света, к окну.
Аналогичным образом насекомые реагируют на разнообразные механические, температурные, химические и оптические раздражения, на силу тяжести, и соответственно у них встречаются во многих различных формах тигмо-, рео-, анемо-, гидро-, гео-, термо-, хемо-, фото- и другие таксисы.
Предметное восприятие
Особый интерес представляет способность насекомых (и других членистоногих) к оптическому восприятию форм как необходимого компонента перцептивной психики. На уровне элементарной сенсорной психики различение форм еще невозможно.
До недавнего времени считалось, что насекомые способны к восприятию формы, но лишь в специфических рамках. Эта специфика усматривалась в том, что в экспериментах насекомые, особенно пчелы, оказывались способными различать лишь фигуры, напоминающие по своим очертаниям цветы (например, кресты или звездчатые фигуры), простые же геометрические фигуры они в этих опытах не различали. Однако Мазохин-Поршняков (в работах, проведенных совместно с Г. М. Вишневской) показал, что пчел можно с полным успехом дрессировать и на такие фигуры, как треугольник, круг, квадрат, из чего он заключил, что пчелы способны распознавать фигуры непосредственно по их графическим признакам.
О способности насекомых к распознаванию форм свидетельствуют и полевые опыты Тинбергена, в которых самки роющих ос обучались распознавать круг из сосновых шишек, выложенных вокруг входа в норку. После того как оса улетала за добычей, круг передвигался на 30 сантиметров в сторону. После возвращения оса искала норку сперва в центре круга (рис. 37). В следующих опытах к тому же шишки заменялись черными камешками, а вокруг норки выстраивался треугольник или даже эллипс из шишек, но оса тем не менее прилетала в круг, хотя из предыдущих опытов было известно, что оса вполне способна отличать камешки от шишек. Таким образом, пространственная ориентация осуществлялась здесь только по форме (кругу).
Хотя у высших насекомых способность к предметному восприятию и не вызывает сомнения, в этом отношении они явно уступают позвоночным; Очевидно, это связано со специфическим строением органов зрения: разрешающая способность глаза зависит у насекомых от количества омматидиев. Чем их больше, тем выше эта способность, так как картина воспринимаемого объекта расчленяется на большее число точек. К тому же насекомые, как и другие членистоногие, очевидно, ориентируются преимущественно все же не по предметным компонентам среды как таковым, а по отдельным их признакам, как это имеет место на стадии элементарной сенсорной психики.
У других беспозвоночных, находящихся также на низшем уровне стадии перцептивной психики, — у головоногих моллюсков — зрение играет не меньшую, если не большую роль, чем у насекомых, и является у них, безусловно, ведущей рецепцией (хотя эти животные ориентируются и хорошо развитыми осязанием и химической чувствительностью). Соответственно и орган зрения получил у головоногих исключительное развитие: головоногие обладают наиболее крупными глазами не только среди обитателей морей, но и вообще в животном мире, причем не только в относительном, но и в абсолютном отношении. Глаза гигантских спрутов (род Architeuthus) достигают в поперечнике 40 сантиметров при общей длине животного (считая и «руки») до 20 метров. Для сравнения можно указать, что у всех, даже самых крупных, китов (например, у 30-метрового голубого кита) диаметр глаза составляет немногим больше десяти сантиметров. Поразительна и огромная разрешающая способность (зоркость) глаза головоногих: на один квадратный миллиметр приходится у разных представителей головоногих от 40 000 до 162 000 палочек, у человека — 120 000–400 000, у совы, обладающей самым зорким глазом в мире, — 680 000. Что же касается предметного восприятия, то головоногие моллюски в отличие от низших моллюсков уже способны к подлинному предметному восприятию, что выражается прежде всего в различении ими формы объектов. Так, осьминог в состоянии отличить треугольник от квадрата, малый квадрат от большого, крест от квадрата, вертикальный прямоугольник от горизонтального, треугольник от ромба и т. д. Всего, как удалось установить, осьминог способен различать 46 разных форм (опыты Б. Б. Бойкотта и Дж. З. Янга). Все же зрение головоногих, хотя и сближается с таковым позвоночных, отличается и рядом примитивных признаков, делающих отражение предметов действительности недостаточно полноценным. Еще предстоит большая исследовательская работа для выяснения специфических особенностей оптического восприятия головоногих.
Общение и групповое поведение
У высших беспозвоночных уже в полном объеме представлена категория поведения, которая, как было показано, только в зачаточной форме обнаруживается у некоторых представителей низших многоклеточных беспозвоночных. Речь идет об общении, которое у высших беспозвоночных проявляется иногда и в очень сложных формах подлинного группового поведения (в отличие от колониальной жизни, свойственной многим низшим беспозвоночным). Напомним, что общение, в точном значении этого термина, появляется только тогда, когда контакты между животными начинают осуществляться посредством специальных сигнальных действий.
У головоногих моллюсков и членистоногих, особенно у насекомых, мы находим четкие системы коммуникации с передачей информации по разным сенсорным каналам. Это относится прежде всего к пчелам и муравьям, что, конечно, связано со сложной, высокодифференцированной структурой их сообществ и разделением функций между членами последних (распределение пищедобывательнои, защитной и воспроизводительной функций между отдельными особями).
У муравьев разные формы химической чувствительности играют, очевидно, большую роль, чем другие виды рецепции, и общение между особями осуществляется прежде всего по этому каналу: личинки муравьев выделяют специфические вещества, побуждающие взрослых рабочих особей кормить их, по запаху обитатели одного муравейника узнают «своих» и «чужих». Даже о том, жив или мертв муравей, его сородичи узнают по специфическим выделениям.
Согласование поведения обитателей муравейника происходит путем подачи и приема химических сигналов. Достаточно указать на химические средства муравьев (а также пчел и ос), выделение которых служит сигналом тревоги. Эти вещества выделяются муравьями в случае опасности и быстро распространяются по воздуху. Их восприятие вызывает у других муравьев («солдат») защитную реакцию (готовность к нападению на врага), хотя источник опасности им еще не известен. Поскольку, однако, радиус действия такого сигнала, подаваемого одним муравьем (например, в случае вторжения в муравейник какого-либо насекомого), невелик, то он способен указать и место, откуда угрожает опасность. Чем больше эта опасность, тем больше муравьев выделяют химические сигналы тревоги и тем больше, следовательно, особей призывается на помощь.
По другим выделениям, оставляемым на субстрате в виде следа, муравьи узнают, куда, в какую сторону ушел их сородич. К тому же интенсивность такого пахучего следа указывает на количество пищи, обнаруженной в том месте, куда ведет след, тем более что, направляясь к муравейнику, муравьи наносили по пути пахучие метки лишь в тех случаях, когда они находили корм. При этом тем больше оставляется меток, чем больше обнаружено корма. Поскольку эти метки довольно быстро испаряются, вновь идущие по этому следу муравьи будут воспринимать лишь свежие метки, а таких будет все меньше по мере приближения кормового объекта. Таким образом, муравьи, в конце концов узнают о том, что пищевые ресурсы в данном месте исчерпаны, и перестанут к нему направляться.
У некоторых пчел также существует химический способ передачи информации о нахождении пищи. Это относится, прежде всего, к пчелам — из подсемейства Meliponinae, у которых нет «языка танца». Эти широко распространенные в тропиках пчелы не имеют жала, но яростно кусаются. У мелких форм (самая маленькая имеет длину всего 1,5 миллиметра) нашедшая пищу сборщица только приводит других пчел в возбужденное состояние, беспокойно бегая по сотам, толкая их и время от времени подбегая к летку. Вылетающие же пчелы ориентируются только по запаху цветов. У других видов возбуждение пчел первоначально достигается подобным же образом, но дополнительно сборщица, принесшая взяток, приводит соты толчками в вибрирующее состояние. После этого она возвращается к обнаруженным цветам, а на обратном пути оставляет на траве, ветках или камнях через каждые два-три метра пахучие метки, по которым затем приводит группами других пчел к источнику пищи. Это достигается тем, что пчела-разведчица летает взад и вперед между двумя соседними метками, пока другие пчелы не присоединяются к ней. Следовательно, здесь мы имеем дело с комплексной передачей информации по оптическому, тактильно-вибрационному и, возможно, акустическому каналам, но при этом ведущую роль играют оставляемые сборщицей запахи.
У нашей медоносной пчелы также существует химический способ передачи информации, но только как вспомогательный, когда она имеет дело с кормом, лишенным собственного заметного запаха. В этих случаях место нахождения кормового объекта метится секретом из так называемой железы Насонова.
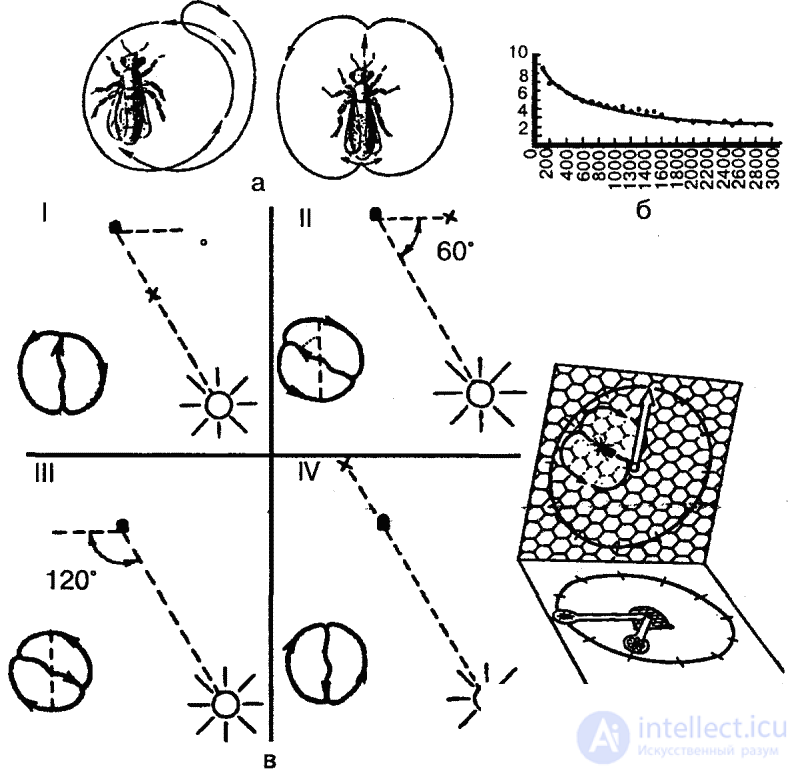
Однако основным и наиболее совершенным способом передачи информации о кормовых объектах являются «танцы». Найдя источник пищи (нектара) и вернувшись в улей, пчела раздает другим пчелам-сборщицам пробы нектара и приступает к «танцу», который состоит из пробежек по сотам. Рисунок танца зависит от местонахождения обнаруженного источника пищи: если он находится рядом с ульем (на расстоянии 2–5 метров от него), то производится «танец-толчок»: пчела беспорядочно бежит по сотам, время от времени виляя брюшком; если же корм обнаружен на расстоянии до 100 метров от улья, то выполняется один из описанных К. Фришем танцев — «круговой», который состоит из пробежек по кругу попеременно по часовой стрелке и против нее (рис. 38). Если же нектар обнаружен на большем расстоянии, то выполняется «виляющий» танец, также описанный и изученный Фришем. Это пробежки по прямой, сопровождающиеся виляющими движениями брюшка с возвращением к исходной точке то слева, то справа (рис. 38, а). Интенсивность виляющих движений указывает на расстояние находки: чем ближе находится кормовой объект, тем интенсивнее выполняется танец (рис. 38, б).
Кроме того, как тоже показал Фриш, во второй форме танца угол между линией пробежки и вертикалью (на вертикально расположенных сотах) соответствует углу между линией полета пчелы от улья к кормовому объекту и положением солнца. Таким образом, горизонтальное направление истинного полета переносится на вертикальную плоскость, ориентация по положению солнца подменяется ориентацией по земному притяжению. При этом, если следует лететь навстречу солнцу, пчела танцует снизу вверх, если же от солнца, то сверху вниз (рис. 38, в). Таким образом, медоносная пчела способна к транспозиции с фотоменотаксисов на геоменотаксисы: фиксированный угол к видимому положению солнца переносится в темном улье к силе тяжести.
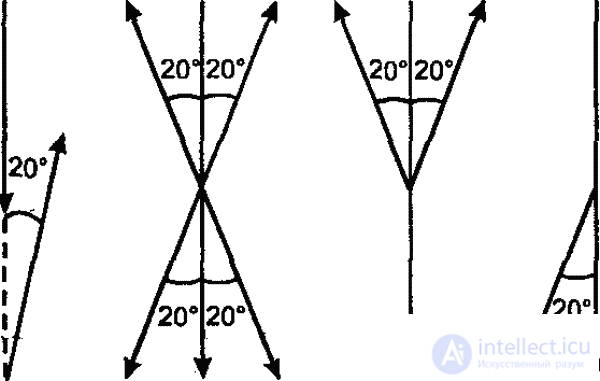
Эта способность медоносной пчелы к транспозиции, коренящаяся в широко распространенной среди насекомых способности к переносу менотаксисных движений на вертикальную плоскость (рис. 39), обеспечивает, как мы видим, исключительную полноту и точность передачи информации. Такая транспозиция является частным случаем трансформации, лежащей в основе всякого общения между животными: свойства действительных предметов и ситуации трансформируются на языке животных с помощью видоспецифического «кода» в оптические, акустические и прочие условные сигналы, информирующие других особей об этих свойствах. Несмотря на всю сложность и точность, система коммуникации медоносной пчелы в принципе не отличается от любой другой системы общения у животных. Переводятся ли истинные пространственные параметры в сигналы, воспринимаемые с помощью зрения, осязания и чувства силы тяжести, как это имеет место у медоносной пчелы, или же в сигналы, воспринимаемые хеморецепцией (муравьи), — информация в любом случае передается в трансформированном, условном виде. Здесь как нельзя нагляднее выступают характер и функциональное значение психики у животных.
Вершиной развития инстинктивных компонентов общения является ритуализация поведения, которая особенно четко выступает в воспроизводительном поведении насекомых, в частности в «ухаживании» самцов за самками. Вот типичный брачный «ритуал» самца бабочки-бархатницы, описанный Тинбергеном: полетев вслед за самкой, самец садится рядом с ней, потом становится перед ней, начинает «дрожать» крыльями (переднезадние движения сомкнутых крыльев), затем повторно раскрывает и закрывает их, что сопровождается постукиванием усиками по крыльям самки. После этого самец обхватывает самку крыльями и зажимает между ними ее усики, которое при этом прикасаются к пахучему участку на крыльях самца. Только после этого, если самка не отвергает «ухаживания» самца, начинается спаривание.
В этом примере обращает на себя внимание строгая фиксированность последовательности сигнальных поз и телодвижений, что и характерно для ритуализованного поведения. Как мы видим, при ритуализации двигательные акты всегда носят условный характер: выполняемые животным движения несут лишь сигнальную, но не непосредственно «механическую» функцию. Именно на этой основе зародился язык поз, телодвижений и звуков у животных во всем своем удивительном многообразии. При этом в условности сигналов находит непосредственное воплощение психический компонент общения.
Наиболее сложные системы общения присущи, конечно, насекомым, живущим большими семьями. Это относится опять-таки в первую очередь к медоносной пчеле и муравьям. Дифференцированные и многообразные взаимоотношения между членами пчелиной семьи, опирающиеся на сложные формы коммуникации, являются уникальными в животном мире. По поводу этого сообщества высказывались самые различные мнения начиная от антропоморфичного толкования ее как копии человеческого общества или государства (сохранилось неудачное, вводящее в заблуждение название «общественные насекомые») до представления о некоей сумме независимо действующих «рефлекторных автоматов». Вместе с тем оказалось (исследования К. П. Трубецкой), что пчелиная семья представляет собой объединение особей, поведение которых носит самостоятельный, но не независимый характер. Самостоятельность поведения отдельных пчел проявляется прежде всего в резко выраженных индивидуальных различиях: предпочтение отдельных видов ульевых работ, различия в сроках и характере перехода от ульевой деятельности к летной и др. Большую сложность в количественном и качественном аспектах достигают взаимоотношения между обитателями улья, в результате чего функциональная настроенность одних пчел может оказать влияние на других. Рабочие особи с их повышенным уровнем нервной деятельности часто сообщают возбуждение другим, менее активным рабочим пчелам.
Территориальное поведение
Уже у кольчатых червей и низших моллюсков наблюдаются первые проявления территориального поведения, что выражается в приуроченности жизнедеятельности к постоянно занимаемому участку, на котором располагается и убежище («дом») животного. Правда, выполнение жизненных функций (или хотя бы части из них) на определенном участке составляет лишь часть территориального поведения, равно как и наличие «дома» само по себе еще не является критерием последнего. При полноценном территориальном поведении, встречающемся лишь на уровне перцептивной психики, животное-хозяин своим видом, голосом или запахом дает знать сородичам о своем присутствии на занятом участке. Важную роль играет при этом «маркировка местности», т. е. нанесение пахучих или иных меток на разные объекты, особенно на периферии участка.
Такими метками могут быть испражнения или выделения кожных желез, а также оптические знаки — содранная со стволов деревьев кора, вытоптанная трава и т. п.
Весьма существенным признаком территориального поведения является активная защита участка хозяином. Тинберген придает этому критерию наряду с привязанностью к местности особое значение. Враждебное отношение («агрессивность») проявляется хозяином по отношению к любому представителю того же вида, особенно того же пола.
Часто такое поведение приурочено лишь к периоду размножения. Так, например, древесный таракан (Cryptocereus punctulatus), устраивающий гнездовую камеру в гнилой древесине, весьма интенсивно (и обычно успешно) защищает место, где находится эта камера, от вторжения других самцов. В случае поражения гнездовую камеру, где находится и самка, занимает победитель.
В высокоразвитых формах территориальное поведение, как показал немецкий этолог А. Хеймер, встречается у стрекоз. Это интересно уже потому, что стрекозы относятся к самым древним насекомым и, появившись в палеозое, дожили до наших дней, не претерпев за эти 50 миллионов лет существенных морфологических изменений. Вместе с тем, как показывает Хеймер, архаичность строения сочетается у них с высокоразвитым поведением, причем не только в сфере территориальности. Этот, казалось бы, парадоксальный факт сохранения древнейших признаков строения при наличии весьма прогрессивных форм поведения вновь подтверждает общее правило несоответствия морфологических и поведенческих признаков, если иметь в виду морфофункциональные отношения на уровне целого организма.
Что же касается территориального поведения стрекоз (речь идет конкретно о равнокрылых стрекозах-красотках Zygoptera), то половозрелые самцы отыскивают места, пригодные для постоянного пребывания, которые и становятся их индивидуальными участками. Эти участки маркируются оптически, а именно путем ежедневных многократных облетов, и энергично обороняются от самцов-сородичей. Внутри индивидуального участка имеются основные и дополнительные места отдыха, а также зона для откладки яиц, границы которой маркируются упомянутым образом. В эту зону самец приводит самку посредством совместно выполняемого с ней «танца» над поверхностью воды. Кроме того, самец «ухаживает» за самкой, выполняя на своей территории особый ритуализованный брачный полет, а во время откладки яиц он охраняет и сторожит самку или соответственно несколько самок, ибо один самец нередко спаривается подряд с несколькими самками. Охрана проявляется в том, что самец отгоняет других мужских особей, пытающихся спариваться с «его» самками, что мешает откладыванию яиц, сторожевая же функция состоит в том, что при попытке самок прервать это занятие самец заставляет их вернуться и возобновить его. Добавим еще, что территориальная борьба самцов выполняется в высокоритуализованных формах (подлинная борьба отсутствует!). В целом все поведение самцов (как и самок) отличается большой пластичностью.
Таким образом, у данных видов стрекоз существует вполне развитое территориальное поведение: занимаются индивидуальные участки, в которых выделяются функциональные зоны (отдыха, размножения), на этих участках выполняются все жизненные функции (за исключением сна: вечером самцы собираются в особых местах ночевок), хозяин дает знать о своем присутствии, маркирует и активно защищает участок.
Интересные особенности территориального поведения муравьев были выявлены советским энтомологом А. А. Захаровым. Оказалось, что у муравьев существуют два основных типа использования кормовых участков: совместное использование угодий несколькими семьями и использование кормового участка населением лишь одного гнезда. При этом выявилась прямая связь между плотностью муравьев на участке и «агрессивностью»: у видов с низкой плотностью участки не охраняются (кроме пригнездовой зоны), при высокой же плотности на кормовых участках появляются охраняемые территории, а между ними «нейтральные зоны» (рис. 40). Как показал другой советский исследователь, Г. М. Длусский, муравьи того же вида, принадлежащие к другим семьям, равно как и представители других видов, в пределы этих территорий не допускаются.
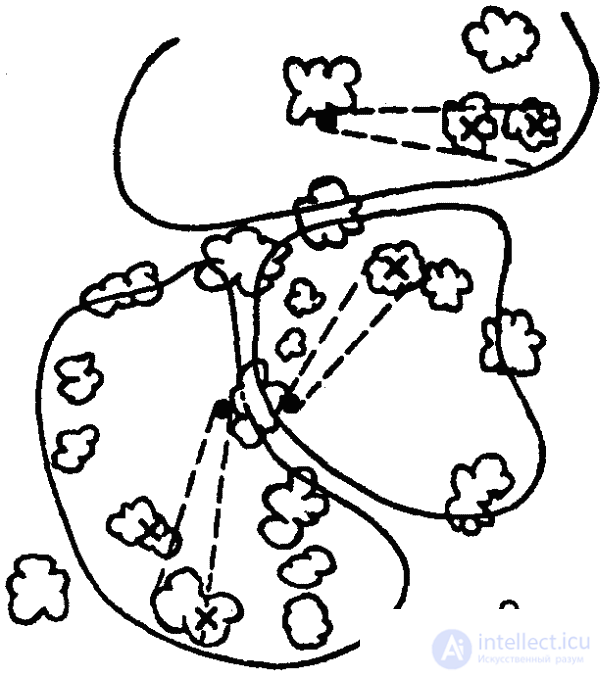
Наибольшую сложность территориальное поведение достигло у рыжих лесных муравьев, у которых наблюдается и наибольшая плотность особей на кормовых участках. Охраняемые территории этих муравьев достигают значительно большей величины, чем у других видов. Центральными элементами такой территории являются муравьиные тропы (постоянные кормовые дороги), длина которых нередко превышает 100 метров, а протяженность всей сети троп — 1 километр. Каждой дорогой пользуется лишь определенная группа муравьев («колонна»), занимающая в муравейнике определенный сектор, примыкающий к «своей» дороге. Территория всей семьи подразделяется дорогами на отдельные части, составляющие территории отдельных колонн. Между ними имеются такие же нейтральные пространства, как и между территориями семей, хотя и не столь выраженные, как в последнем случае. Границы территории маркированы и препятствуют «рассеиванию» муравьев. Муравьи двигаются по территории не хаотично, а в соответствии со структурой территории: в центральной и средней ее частях — по кратчайшему пути от дорог или муравейника, на периферии же — параллельно границам территории. Таким образом, траектория движения каждого муравья детерминируется местом нахождения его на территории (исследования И. В. Стебаева). Наибольшую охотничью активность муравьи развивают на периферии территории (далее 10 метров от гнезда). Сбор строительного материала производится в пределах двухметровой полосы вдоль дорог.
Захаров справедливо отмечает сходство территориального поведения муравьев с таковым у птиц и хищных млекопитающих.
Инстинкт и научение в поведении насекомых
Долгие годы господствовало мнение будто насекомые и другие членистоногие являются существами, поведением которых руководит жесткий «слепой инстинкт». Это представление укоренилось преимущественно под влиянием работ выдающегося французского энтомолога Ж. А. Фабра, который сумел своими блестящими исследованиями убедительно показать, что даже сложнейшие действия насекомых не являются проявлением «разума», а выполняются на врожденной, инстинктивной основе. Об этом говорит сайт https://intellect.icu . Одностороннее развитие положений Фабра и привело к указанной, неверной оценке поведения насекомых, к отрицанию не только разумности их поведения, но и к отрицанию, или хотя бы умалению роли накопления индивидуального опыта, научения в их жизни.
Как мы уже видели, формирование любой формы видотипичного, наследственно «закодированного», т. е. инстинктивного, поведения в онтогенезе всегда сопряжено в той или иной степени с какими-либо элементами индивидуально приобретаемого поведения, научения. О строго фиксированном инстинктивном поведении в «чистом виде» не приходится говорить даже относительно низших животных.
В полной мере это относится и к насекомым, инстинктивное поведение которых также совершенствуется научением. В этом состоит основная роль научения в жизни насекомых. Можно, очевидно, считать, что научение стоит у насекомых и других членистоногих «на службе» у инстинктивного поведения. Как и у других животных, инстинктивные движения (врожденные двигательные координации) у них генетически строго фиксированы. Инстинктивные же действия, инстинктивное поведение являются и у насекомых в той или иной степени пластичными благодаря включению в них благоприобретаемых компонентов.
В естественных условиях способность к накоплению индивидуального опыта проявляется у насекомых в неодинаковой степени в разных функциональных сферах. Чаще всего она связана с ориентацией в пространстве и пищедобывательной деятельностью. Примером могут служить отмеченные выше опыты по обучению пчел за пищевое подкрепление ориентироваться по различным рисункам. Другой пример — муравьи, которые очень легко (всего за 12–15 опытов) научаются проходить даже сложный лабиринт, но, насколько известно, не научаются действиям, лежащим за пределами указанных функциональных сфер. Такая специфическая направленность (и одновременно ограниченность) способности к научению является характерной особенностью научения у представителей всего типа членистоногих.
Роль научения в поведении насекомых наглядно выступает и в «танцах» пчел — этих высших представителей членистоногих. Отстаивая взгляд, будто насекомые, в том числе пчелы, являются «стимульно связанными, рефлекторными животными», американские ученые В. Детьер и Э. Стеллар заявляют, например, что выполнению и интерпретации сложного танца пчелы не обучаются. Вместе с тем, как показали советские исследователи Н. Г. Лопатина, И. А. Никитина, Е. Г. Чеснокова и другие, процессы научения не только уточняют, но и модифицируют коммуникационные способности пчелы в онтогенезе и расширяют набор сигнальных средств.
Более того, как установили названные исследователи, биологическая значимость сигнальной деятельности медоносных пчел определяется стереотипом условных рефлексов, приобретаемых в онтогенезе по мере освоения пространства и при общении в семье. Оказалось, что интерпретация передаваемой в танце информации о расстоянии и направлении полета к источнику пищи возможна лишь в том случае, если пчела до этого научилась соотносить местонахождение корма с характером информации, содержащейся в танце сборщиц. Кроме того, тактильный компонент танца (вибрации брюшка) не имеет врожденного сигнального значения. Последнее приобретается в онтогенезе также условно-рефлекторным путем: пчелы, не имевшие в онтогенезе контактов (пищевых) с танцовщицей, не в состоянии интерпретировать этот существеннейший элемент танца. Следовательно, каждая пчела должна в основном научиться «понимать» язык танца. С другой стороны, образование временных связей оказалось важным и для формирования самой способности к выполнению танцев.
Таким образом, нет неизменных форм поведения даже там, где прежде всего требуется стереотипность, — в сигнальных позах и телодвижениях. Даже такое врожденное коммуникативное поведение, как «танцы» пчел, не только дополняется и обогащается процессами научения, не только переплетается с ними, но и формируется в комплексе с индивидуально приобретаемыми элементами поведения.
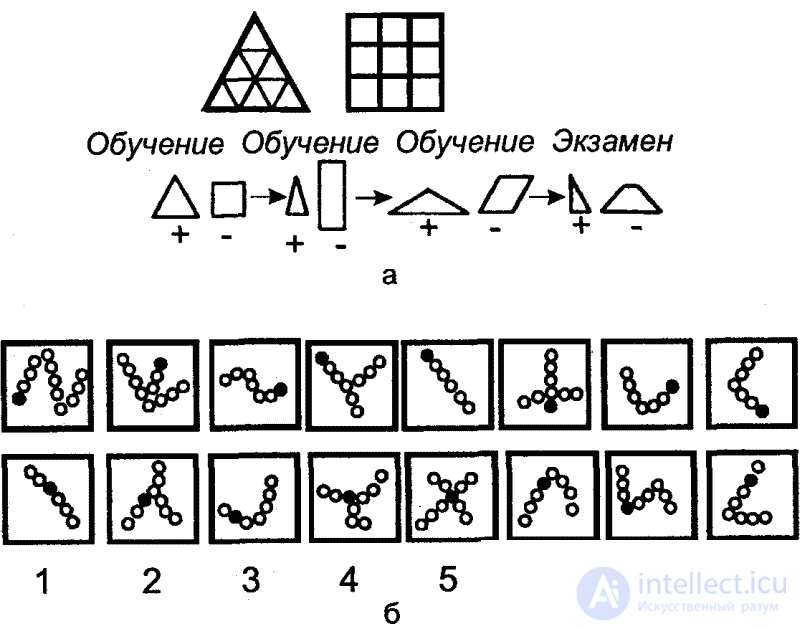
Конечно, медоносная пчела занимает среди насекомых исключительное положение, и далеко не у всех представителей этого огромного класса психическое развитие достигает такой высоты. Об исключительных психических качествах медоносной пчелы свидетельствуют, в частности, экспериментальные данные, говорящие о наличии у нее аналогов некоторых психических функций высших позвоночных. Речь идет об установленной Мазохиным-Поршняковым высокоразвитой способности пчелы к зрительным обобщениям, например, типа «треугольник» и «четырехугольник» (независимо от конкретной формы, соотношения размеров и взаимной ориентации фигур) (рис. 41, а), «двуцветность» и др. В одной из серий опытов пчелам предлагалось выбрать из попарно предъявляемых фигур те, у которых один локальный признак (зачерченный кружочек) находился на конце цепочки из кружочков независимо от длины и формы этих цепей (рис. 41, б). Со всеми предложенными им задачами, даже в наиболее сложных вариантах, пчелы вполне справлялись. При этом отмечалась большая пластичность, нестандартность поведения, что экспериментатор справедливо связывает с непрерывной изменчивостью условий среды (непостоянство освещения, взаимного расположения, формы, окраски и многих других признаков компонентов среды), при которых этим насекомым приходится добывать пищу. Мазохин-Поршняков приходит к выводу, что выбор незнакомого объекта на основе обобщенных зрительных образов (иногда неправильно обозначаемых им как «понятия») есть свидетельство нестандартного использования пчелами индивидуального опыта, его применения в новой ситуации, отличной от обстановки первоначальной выработки соответствующего навыка.
Таким образом, здесь справедливо подчеркивается наличие и значение факта переноса определенного навыка в новую ситуацию и решение сложной задачи на основе индивидуального опыта, зафиксированного в виде обобщенного зрительного представления. В этом отношении мы действительно находим уже у пчел психические способности, аналогичные тем, которые относятся к предпосылкам интеллектуальных действий высших позвоночных животных. Однако одних этих предпосылок недостаточно для интеллектуального поведения, мышления животных, особенно если смотреть на эти высшие психические функции животных как на ступень по направлению к зарождению человеческого сознания. Поэтому описанные способности пчел не могут служить и критерием для признания у них мышления и уж во всяком случае не приходится говорить о наличии у пчелы рассудочной деятельности, хотя бы и в элементарной форме, как толкует результаты своих исследований Мазохин-Поршняков. Признавая у высших животных наличие своеобразных мыслительных способностей, интеллекта, нужно со всей определенностью отдавать себе отчет в том, что рассудок, т. е. разум, сознание как качественно иная категория психического отражения не присущи ни одному из животных, а только человеку.
Общая характеристика низшего уровня перцептивной психики
На низшем уровне перцептивной психики уже представлены все те прогрессивные признаки, которые характеризуют перцептивную психику вообще, но во многих отношениях поведение относящихся сюда животных носит и примитивные черты, сближающие его с поведением нижестоящих животных. Так, основную роль играет ориентация поведения по-прежнему по отдельным свойствам предметов, но не по предметам как таковым: предметное восприятие явно играет еще подчиненную роль в общем поведении. Равным образом в последнем преобладают ригидные, «жестко запрограммированные» элементы поведения над гибкими, благоприобретаемыми и т. д.
С другой стороны, мы констатируем на этом уровне четко выраженный активный поиск положительных раздражителей, т. е. положительное таксисное поведение получает мощное развитие. Налицо все виды высших таксисов, включая мнемотаксисы. Последние играют в пространственной ориентации особенно существенную роль, и именно в индивидуальном заучивании ориентиров проявляется в наибольшей степени способность к благоприобретаемому изменению видового поведения, к научению.
Вместе с тем важно подчеркнуть, что хотя у рассматриваемых здесь животных, в частности насекомых, накопление индивидуального опыта, научение играют существенную роль, наблюдается и определенная противоречивость в процессах научения, сочетания прогрессивных и примитивных черт. Специфическая направленность, приуроченность этих процессов к определенным функциональным сферам, как и само подчиненное положение, которое занимает научение по отношению к инстинктивному поведению, несомненно, указывают на переходное положение данного уровня психического развития между элементарной сенсорной и развитой перцептивной психикой.
Но, как мы видели, это никоим образом не означает, что насекомым, как и другим представителям рассматриваемой группы животных, недостает пластичности поведения. Наоборот, и здесь в полной мере проявляется общая закономерность, что усложнение инстинктивного поведения неизбежно сочетается с усложнением процессов научения (и наоборот). Только такое сочетание обеспечивает подлинный прогресс психической деятельности.
Инстинктивное поведение представлено на рассматриваемом уровне психического развития уже весьма развитыми новыми категориями: групповое поведение, общение, ритуализация. Особую сложность приобретают формы общения у видов, живущих огромными семьями, из которых лучше всего изучены пчелы. Язык пчел, этих высших представителей членистоногих, относится к наиболее сложным формам общения, которые вообще существуют в животном мире. Возможно, что новые исследования познакомят нас с поразительными психическими способностями и других насекомых, но на сегодняшний день пчелы представляются нам наиболее развитыми в этом отношении. Наиболее сложные формы инстинктивного поведения закономерно сочетаются у них с наиболее разнообразными и сложными проявлениями научения, что обеспечивает не только исключительную согласованность действий всех членов пчелиной семьи, но и максимальную пластичность поведения особи. Психические способности пчел (как и некоторых других высших насекомых) в некоторых отношениях, очевидно, уже выходят за рамки низшего уровня перцептивной психики.
В ином направлении, чем у членистоногих, шло развитие психической активности у головоногих моллюсков. По некоторым признакам они приблизились к ветви, ведущей к позвоночным, о чем уже свидетельствуют их крупные размеры и отмеченные выше особенности строения нервной системы и особенно зрительного рецептора, что непосредственно связано с резким увеличением скорости движения по сравнению с другими моллюсками.
Поведение головоногих еще совершенно недостаточно изучено, но уже известны многие примечательные их способности. Прежде всего они выделяются существенным усложнением инстинктивного поведения. У головоногих уже встречаются территориальное поведение (занятие и защита индивидуальных участков), «агрессивность», которая только намечается у высших червей, групповое поведение (стайная жизнь кальмаров и каракатиц), в сфере размножения появляются ритуализованные формы поведения, что находит свое воплощение в видоспецифическом «ухаживании» самцов за самками. Все это присуще только высшим животным, помимо головоногих, членистоногим и позвоночным.
Рядом исследователей обращалось особое внимание на весьма развитое у осьминогов «любопытство», что выражается в обследовании ими биологически «бесполезных» предметов, а также на их высокоразвитые манипуляционные и конструктивные способности. Эти способности проявляются в строительстве валов и убежищ из камней, панцирей крабов, раковин устриц и т. п. Этот строительный материал осьминог подбирает, переносит и укрепляет «руками». Иногда такие гнезда представляют собой закрытые со всех сторон строения. По некоторым, правда еще не подтвержденным, наблюдениям, осьминоги способны и к орудийным действиям, пользуясь камнями для защиты.
Очень важным представляется и то обстоятельство, что впервые у головоногих появляется способность к установлению контактов с человеком, к общению с ним, результатом чего является возможность подлинного приручения этих животных (в отличие от насекомых!).
Таким образом, головоногие, оставив далеко позади других моллюсков, как и вообще всех низших беспозвоночных, достигли, несомненно, высокого уровня психического развития, во многом сблизившись с позвоночными животными.
Вместе с тем и у головоногих наблюдается та же противоречивость в способности к научению, что и у насекомых. Так, например, у осьминога вообще хорошо развита способность к научению на зрительные и тактильные стимулы, но в ряде случаев он оказывается не в состоянии решить, казалось бы, несложные задачи. Особенно это относится к преодолению преград: осьминог неспособен находить обходной путь, если приманка (краб) располагается за прозрачной преградой (в стеклянном цилиндре или за проволочной сеткой); Тщетно пытаясь овладеть приманкой в прямом направлении, осьминог не хватает ее сверху, через край. Правда, по Бойтендайку, некоторые осьминоги все же способны решить несложные задачи обходного пути. При этом, очевидно, большое значение имеет прежний опыт особи. Другие головоногие уступают осьминогу по своим психическим способностям.
Конечно, при оценке подобных экспериментов необходимо иметь в виду, что здесь ставятся биологически неадекватные, а поэтому и неразрешимые задачи: в естественных условиях осьминог никогда не оказывается в ситуации, когда непосредственно зримая жертва оказывается недосягаемой. К тому же задачи обходного пути относятся к разряду весьма сложных — с ними не справляются не только черепахи, но и куры (в биологически несравненно более адекватных условиях). Тем не менее следует думать, что в психической деятельности головоногих действительно сочетаются прогрессивные черты, сближающие их с позвоночными, с примитивными — наследством низших моллюсков. К примитивным чертам относится и известный «негативизм» научения: головоногие легче научаются избегать неприятных раздражений, чем находить благоприятные. И в этом нетрудно усмотреть общность с поведением животных, обладающих элементарной сенсорной психикой.
На низшем уровне перцептивной психики находится, очевидно, хотя бы отчасти и ряд представителей низших позвоночных. Однако в корне различные строение и образ жизни членистоногих и позвоночных являются причиной того, что и их поведение и психика, в сущности, несопоставимы. Так, одной из отличительных особенностей насекомых являются их малые по сравнению с позвоночными размеры. В связи с этим окружающий насекомое мир представляет собой нечто совершенно особое: это не микромир простейших, но и не макромир позвоночных. Человеку трудно себе представить этот мир насекомых с его (с нашей точки зрения) микроландшафтами, микроклиматами и т. д. Хотя насекомые живут рядом и вместе с нами, они живут в совершенно иных условиях температуры, освещения и т. п. Уже поэтому психическое отражение действительности не может у насекомых не быть принципиально иным, чем у позвоночных, да и у большинства других беспозвоночных.
Но поскольку наиболее общие признаки психического отражения, характерные для данного его уровня, присущи всем упомянутым животным, можно, очевидно, по поводу рассмотренных нами в качестве примера насекомых сказать, что мы имеем здесь дело с типичными проявлениями низшего уровня перцептивной психики, но в формах, отвечающих тем особым условиям жизни этих животных, которые отмечались выше.
Высший уровень развития перцептивной психики
В мире животных процесс эволюции привел к трем вершинам: позвоночные, насекомые и головоногие моллюски. Соответственно высокому уровню строения и жизнедеятельности этих животных мы наблюдаем у них и наиболее сложные формы поведения и психического отражения. Представители всех трех «вершин» способны к предметному восприятию, хотя, очевидно, только у позвоночных эта способность получила полное развитие. У остальных двух групп перцепция развивалась своеобразными путями и качественно отличается от таковой у позвоночных. Аналогично обстоит дело и с другими решающими критериями стадии перцептивной психики, не говоря уже о том, что высшего уровня перцептивной психики достигли в процессе эволюции вообще только представители позвоночных, и то явно не все. Только у высших позвоночных обнаруживаются все наиболее сложные проявления психической деятельности, которые вообще встречаются в мире животных.
Сравнивая беспозвоночных с позвоночными, необходимо также учесть, что ни головоногие, ни членистоногие не имеют никакого отношения к предкам позвоночных. Путь, ведущий к этим вершинам, отклонился от пути к третьей вершине еще на очень раннем этапе эволюции животного мира. Поэтому высокое развитие морфологических и поведенческих признаков, отмеченных нами у этих животных, является при сравнении с позвоночными лишь аналогией, обусловленной мощным повышением общего уровня жизнедеятельности, характерным для всех трех групп животных.
С филогенетической точки зрения для нас значительно больший интерес могли бы представить иглокожие, которые так же, как и позвоночные, относятся к вторичноротым в отличие от первичноротых, у которых центральная нервная система располагается на брюшной стороне тела и к которым, в частности, относятся моллюски и членистоногие. Однако и современные иглокожие (морские ежи, звезды и т. д.) являются не предками позвоночных, а лишь боковой ветвью представленной весьма специализированными формами с примитивным поведением, характерным для элементарной сенсорной психики.
На этом уровне находятся и низшие хордовые, которые вместе с позвоночными (или «черепными») составляют тип хордовых. К низшим хордовым относятся оболочники и бесчерепные. Оболочники — морские животные, часть которых ведет неподвижную жизнь (асуи-дия). Бесчерепные представлены всего двумя семействами с тремя родами мелких морских животных, наиболее известное из которых — ланцетник. У бесчерепных и позвоночных общим признаком является внутренний осевой скелет, имеющий вид сплошного (хорда ланцетника) или членистого (позвоночник) стержня, над которым располагается центральная нервная система, имеющая форму трубки. Позвоночные подразделяются на классы круглоротых (миноги и миксины), рыб, земноводных, пресмыкающихся, птиц, млекопитающих. К высшим позвоночным относятся только последние два класса, в пределах которых, следовательно, и обнаруживаются проявления высших психических способностей животных. Что же касается низших позвоночных, то здесь намечаются промежуточные ступени психического развития, характеризующиеся разными сочетаниями элементов низшего и высшего уровней перцептивной психики.
У позвоночных нервная трубка образует в головном отделе вздутия, превращающиеся в ходе эмбриогенеза в головной мозг. Уже у наиболее примитивных позвоночных, у круглоротых, имеются все пять отделов головного мозга (продолговатый, задний, средний, промежуточный и передний мозг). Процесс дифференциации и прогрессивного развития мозговых структур достигает, как известно, своей вершины у млекопитающих, причем не только в переднем мозгу (большие полушария и их кора), но и в стволовой части головного мозга, где формируются, в частности, центры высших форм инстинктивного поведения.
Что касается млекопитающих, то они вообще занимают особое место в эволюции животного мира, ибо, как указывал Северцов, «млекопитающие очень редко приспособляются к быстро наступающим переменам в окружающей среде (например, к новым врагам, к новой добыче и т. д.) обычным путем, то есть путем медленного изменения своих органов и их функций. Гораздо чаще это происходит путем быстрого изменения прежних привычек и навыков и образования новых, приспособленных к новым условиям среды. Здесь впервые выступает на сцену совершенно новый и необычайно важный фактор адаптивной эволюции позвоночных животных, а именно их психика».[53]
В первой и второй частях этой книги рассматривалось преимущественно поведение высших позвоночных. По этой причине мы ограничимся здесь лишь некоторыми дополнениями к сказанному.
Локомоция
Мышечная система позвоночных состоит из симметрично расположенных вдоль тела сегментов. Правда, посегментное расположение мышц выражено у хордовых слабее, чем у членистоногих и червей, а у высших представителей позвоночных оно сильно нарушено. Различают соматическую мускулатуру, обслуживающую эффекторные органы, и висцеральную — мускулатуру внутренних органов и кожи. Соматическая мускулатура состоит всегда из поперечно-полосатых мышечных волокон.
Конечности позвоночных представлены непарными и парными образованиями. Непарные конечности имеются только у круглоротых и рыб. Это спинной, заднепроходный и хвостовой плавники. Парные конечности, передние и задние, сильно различаются у разных позвоночных соответственно своей функции (плавники, крылья, лапы, ласты, ноги, руки), однако сравнительное изучение строения их скелета позволяет отчетливо проследить эволюционные преобразования от общей примитивной начальной формы.
Основная функция конечностей состоит у всех животных в локомоции, в перемещении животного в пространстве. Однако у многих членистоногих и позвоночных сюда добавляется еще функция опоры приподнятого над субстратом тела. Поэтому в этих случаях говорят об опорно-локомоторной функции конечностей. Не вдаваясь здесь в детальное рассмотрение разнообразных форм этой основной функции конечностей позвоночных, укажем лишь на некоторые существенные моменты.
Видный советский ученый Н. А. Бернштейн писал, что постепенно назревшая в филогенезе потребность в быстрых и мощных движениях — привела на одной из его ступеней к возникновению и параллельному развитию «костно-суставных кинематических цепей скелета» и поперечнополосатой мускулатуры, снабженной соответственными нервными образованиями. У позвоночных эти скелетно-мышечные системы («неокинетические системы», по Бернштейну) получают существенное прогрессивное развитие по сравнению с членистоногими, причем особенно это относится к рассматриваемым здесь высшим позвоночным, в частности к их локомоторным способностям. Бернштейн указывает в этом отношении на глубокие качественные различия между низшими и высшими позвоночными, обусловленные усложнением возникающих перед организмом двигательных задач, возрастанием разнообразия реакций, требующихся от организма, более высокими требованиями в отношении дифференцированности и точности движений. «Достаточно напомнить, — пишет Бернштейн, — насколько, например, аэродинамический полет птицы сложнее почти полностью гидростатического плавания рыбы или насколько богаче по контингентам участвующих движений охота хищного млекопитающего по сравнению с охотой акулы. Молодая отрасль проворных теплокровных млекопитающих победила тугоподвижных юрских завров именно своей более совершенной моторикой».[54]
В современных этологических исследованиях локомоторная активность изучается в ее видотипичных проявлениях как адаптация к специфическим условиям существования: разновидности и особенности ходьбы, бега, прыганья, лазанья, плавания, полета и т. д. определяются особенностями образа жизни и служат важным приспособлением к окружающей среде. Вместе с тем всем формам локомоции присуща ритмичность, выражающаяся в том, что движения выполняются в четкой последовательности многократно и относительно стереотипным образом (хотя поведение животного в целом при этом не является стереотипным). Эта ритмичность основана на эндогенной центрально-нервной стимуляции и проприоцепторной обратной связи. Помимо проприоцепторной чувствительности внешние импульсы лишь регулируют эти ритмы, соотносят их параметры (сила, скорость, длительность движений и т. д.) с конкретными условиями тех ситуаций, в которых оказывается животное. В частности, внешние раздражения вызывают начало или конец локомоторных движений, хотя и это может произойти в результате эндогенной стимуляции.
Сказанное достаточно поясняет тот факт, что локомоторные движения относятся к наиболее «автоматизированным» и однообразно выполняемым компонентам всей двигательной сферы животных. В этой же связи стоит и относительная малочисленность форм локомоции у каждого вида. Определяющей для локомоции является ее физическая, механическая функция. Сами локомоторные движения дают животному только минимальную информацию об окружающем мире.
При этом необходимо, правда, учесть, что локомоторная активность включает в себя и ориентировочные компоненты, имеющие, конечно, и определенное познавательнее значение. Так, например, прыгающие животные, особенно древесные, должны перед прыжком точно «рассчитать» расстояние. Как показали советские исследователи поведения животных В. М. Смирин и О. Ю. Орлов, это делается с помощью особых движений «взятия параллакса» (рис. 42). Оказавшись в новом месте, летяга «прицеливается» к разным предметам, это же она делает перед каждым прыжком, хотя со временем число таких движений уменьшается. В итоге уходящее от опасности животное придерживается заранее «отработанного» пути без лишних движений и совершает прыжки с поразительной точностью.

Манипулирование
Эффекторные органы позвоночных всегда выполняют сразу несколько функций — помимо основной функции еще большее или меньшее число дополнительных. В этом проявляется мультифункциональность этих органов. Что касается конечностей высших позвоночных, то, как уже отмечалось, их основой является опорно-локомоторная функция: все же дополнительные их функции сводятся к разнообразным формам манипулирования предметами (пищевыми или несъедобными). Особый интерес представляют для зоопсихолога формы манипулирования передними конечностями, которые в процессе эволюции привели к орудийной деятельности приматов и тем самым стали важнейшей биологической предпосылкой зарождения трудовых действий у древнейших людей.
Говоря о манипуляциях, дополнительных функциях передних конечностей у высших позвоночных, необходимо иметь в виду, что сложные и разнообразные дополнительные функции присущи у этих животных и ротовому аппарату, причем существует далеко идущее функциональное взаимодействие между этими двумя основными эффекторными системами. Поэтому целесообразно анализировать дополнительные функции передних конечностей и ротового аппарата в комплексе. Так, например, у взрослых лисиц обнаруживается 45 дополнительных функций передних конечностей и ротового аппарата, у барсука — 50, у енота — около 80, а у низших обезьян (макаков и павианов) — свыше 150 (данные Фабри).
Важно отметить, что по мере специализации млекопитающих в сфере локомоции часть дополнительных функций передних конечностей передается челюстному аппарату. Так, например, в ряду медведь — енот — барсук — лисица конечности лисицы в наибольшей степени приспособлены к продолжительному быстрому бегу. Но при этом дополнительные (манипуляторные) функции конечностей сводятся к минимуму и соответствующие жизненно необходимые движения (например, в сфере питания) выполняются почти исключительно ротовым аппаратом. Как раз наоборот обстоит дело у медведей.
Если иметь в виду познавательное значение манипулирования, то следует особенно выделить те его формы, при которых животное удерживает предмет перед собой свободно на весу обеими или одной конечностью. Такая фиксация предмета дает животному наилучшие возможности для комплексного (оптического, ольфакторного, тактильного, кинестетического и пр.) обследования, особенно в ходе деструктивного воздействия на него (практического анализа). По данным, полученным Фабри, это дает при известных условиях обезьянам, а отчасти и другим способным на это млекопитающим наиболее всестороннюю и разнообразную информацию, необходимую для развития высших форм психической активности. Оказалось, что медведи владеют тремя способами фиксации объекта на весу, еноты — шестью, низшие обезьяны и полуобезьяны — тремя десятками таких способов! К тому же только обезьяны обладают достаточно различными двигательными возможностями, чтобы произвести подлинный деструктивный анализ (расчленение) объекта на весу.
О формировании манипуляционной активности уже говорилось раньше (см. ч. II). Напомним лишь, что первым начинает функционировать челюстной аппарат, позже появляются движения передних конечностей, которые, однако, вначале еще нуждаются в поддержке более сильной системы челюстного аппарата. Впоследствии движения конечностей становятся все более сильными и самостоятельными и одновременно все более разнообразными, т. е. возрастает их мультифункциональность. Ряд функций, в частности хватательные, первоначально выполнявшиеся челюстями, переходит при этом к конечностям. Вершиной этого развития являются дифференцированные движения, выполняемые только одной конечностью. Как уже отмечалось, прогрессивное развитие дополнительных двигательных функций передних конечностей являлось и является важнейшим фактором психического развития.
Комфортное поведение. Сон и покой
Разновидностью манипулирования можно отчасти считать комфортное поведение, служащее уходу за телом животного, с той лишь особенностью, что объектом манипулирования является не посторонний предмет, а именно собственное тело. Но, кроме того, сюда относятся и нелокализованные движения, производимые всем телом и лишенные специальной пространственной направленности. В итоге можно выделить следующие категории комфортных движений: очищение тела, потряхивание, почесывание (определенного участка тела об субстрат), катание по субстрату, купание (в воде, песке и т. д.).
Комфортное поведение широко распространено и среди членистоногих (достаточно вспомнить, как муха чистит лапками голову, крылья и т. п.), иногда встречаются и специальные морфологические образования или даже органы для очищения тела. Инстинктивная природа комфортных движений обнаруживается в видоспецифичности их выполнения, равно как в строгой фиксированности участков тела, очищаемых тем или иным органом и способом. Столь же четко видотипичными являются и позы сна и покоя. Так, например, у зубров, бизонов и их гибридов было описано всего 107 видотипичных поз и телодвижений по 8 сферам поведения (исследования М. А. Дерягиной). Наибольшее число поз (2/3) приходится на сферы сна, покоя и комфорта. При этом любопытно, что у детенышей (до 2–3 месяцев) всех трех изученных групп животных видотипичные различия в этих сферах еще не выражены. Напрашивается вывод, что эти различия формируются постепенно в более позднем возрасте.
Если рассмотреть одну из форм комфортного поведения, облизывание, то, согласно Дерягиной, оказывается, что только бизон облизывает стоя корень хвоста или приподнятую переднюю ногу, но только зубр облизывает стоя бедра задней ноги и т. д. Если не считать гибридов, то существует несколько способов облизывания, из них присущих только зубру — один, только бизону — четыре, общих зубру и бизону — четыре. Здесь опять проявляются четкие различия в инстинктивных движениях у близкородственных видов (зубр и бизон рассматриваются иногда даже как подвиды).
Сенсорные особенности
В полном соответствии с высокоразвитыми эффекторными системами высших позвоночных находится и высокий уровень их сенсорных способностей. Наибольшее значение имеют органы слуха и равновесия (внутреннее ухо, начиная с земноводных — дополнительно среднее ухо, а у млекопитающих также и наружное ухо), обоняния (обонятельные мешки и раковины) и зрения (глазные яблоки). Хорошо развиты также кожная и мышечная (тактильно-кинестетическая) и термическая чувствительность, вкус, а в ряде случаев и другие виды чувствительности (электрическая, вибрационная и др.). Соответственно развиты у позвоночных и разные таксисы.
Сопоставление роли зрения и обоняния в жизни высших позвоночных показывает, что у большинства млекопитающих ведущую роль играет обоняние, которое достигает у них удивительной остроты (только акулы могут в этом отношении сравниваться с ними). Так, крот чувствует добычу сквозь толщу земли, белый медведь — сквозь лед. Бурый медведь чувствует на расстоянии 20 метров мед, зарытый в землю на глубину 50 сантиметров. Хорошо известны поразительные обонятельные способности собак и копытных.
Однако, как уже отмечалось, в каждом конкретном случае степень развития обоняния (и соответствующих морфологических структур в рецепторе и головном мозге) определяется особенностями биологии вида. Так, например, по Б. С. Матвееву, даже у родственных видов, обитающих в сходных условиях, но различающихся по биологии, — у обыкновенной полевки (Mictorus arvalis) и полевой мыши (Apodemus agrarius) — наблюдаются существенные сенсорные различия. Полевые мыши поедают разнообразный корм — много семян, а также животный корм, полевки же — преимущественно зеленый корм, который всегда имеется в избытке. Вследствие этого обоняние развито у полевой мыши сильнее, чем у полевки.
Зрение лучше всего развито у птиц и приматов. Оно играет важную роль в их пищедобывательной, оборонительной, воспроизводительной и других формах поведения, обеспечивая четкую ориентацию животного в пространстве. Особое значение приобретает зрение при дальней ориентации мигрирующих птиц, а также внутритерриториальной ориентации млекопитающих. Хорошо изучена, например, ориентация птиц по топографическим признакам (например, береговой линии), поляризованному освещению небосвода и астрономическим ориентирам — солнцу, звездам. Последний случай представляет собой наглядный пример ориентации на основе менотаксисов.
У многих млекопитающих ориентация на своей территории (индивидуальном участке или территории стада) осуществляется с помощью фото- и хемомнемотаксисов (по оптическим и ольфакторным меткам). Кроме того, даже у хуже видящих млекопитающих (например, куньих) зрение играет немаловажную роль при ориентации по подвижным объектам, особенно во время ловли добычи. Даже животные с определенно плохим зрением, как, например, моржи, которые над водой близоруки, пытаются ориентироваться с помощью не только обоняния, но и зрения, особенно опять-таки когда имеют дело с подвижными объектами (приближающимися к ним животными или людьми).
Наиболее мощным зрением, очевидно, во всем мире животных обладают хищные птицы. Достаточно сказать, что сокол способен при благоприятных условиях увидеть сидящего голубя на расстоянии почти полутора километров. Равным образом грифы находят свой корм (трупы животных) на огромном удалении с помощью зрения. Но они не в состоянии обнаружить прикрытые останки животных. Но это относится лишь к птицам, обитающим в степных просторах Восточного полушария. Грифы же Центральной и Южной Америки (например, черные американские грифы), разыскивающие пищу в густых лесах, не могут пользоваться зрением и находят ее, как это ни кажется невероятным, не менее успешно и с не меньшего расстояния с помощью обоняния. Аналогично обстоит дело и у млекопитающих. У копытных, обитающих на открытых пространствах, сильнее развито зрение, чем у их родственников, живущих в лесах, даже если животные принадлежат к одному и тому же роду (например, бизон — преимущественно степное животное и зубр — обитатель лесов).
Все эти примеры наглядно показывают, что образ жизни, биология всецело определяют характер поведения и психики животных (в отличие от человека, который благодаря своей общественно-трудовой деятельности достиг далеко идущей независимости от биологических факторов и психическая деятельность которого обусловливается социальными условиями его жизни). В приведенных примерax это положение материалистической зоопсихологии относится конкретно к сенсорной сфере животных, в частности к вопросу о ведущей рецепции. Как мы видели, в каждом отдельном случае, даже у близкородственных видов, она определяется конкретными условиями жизни животного. Но то же самое относится и к психическому отражению в целом, характер которого также всегда определяется этими условиями.
Зрительные обобщения и представления
Подлинная рецепция, истинное восприятие предметных компонентов среды как таковых возможны лишь на основе достаточно развитой способности к анализу и обобщению, ибо только это позволяет полноценно узнавать постоянно меняющие свой вид (и другие свойства) предметные компоненты среды.
Исследования, проведенные на разных видах позвоночных (помимо круглоротых), показали, что все они способны к предметному восприятию, в частности к восприятию форм. Уже на этом основании можно заключить, что все позвоночные находятся на стадии перцептивной психики. Однако внутри этой стадии наблюдаются существенные различия между низшими и высшими позвоночными.
Так, рыбы способны в эксперименте отличать геометрические фигуры, например квадрат от треугольника, независимо от меняющейся величины этих фигур. Но достаточно в контрольном опыте перевернуть треугольник (острием вниз) или квадрат (придать ему положение ромба), как рыба перестает узнавать эти фигуры. Следовательно, восприятие и узнавание формы оказываются здесь ограниченными, недостаточно гибкими. Отсутствует способность к быстрому переносу сформировавшегося зрительного представления на основе обобщения. Млекопитающие же вполне способны к такому обобщению и легко узнают треугольник или другую геометрическую фигуру любой величины в любом положении. Существенно превосходят рыб также птицы. Правда, и курица, по некоторым данным, не узнает перевернутый треугольник, который до этого безошибочно отличала от другой геометрической фигуры. Вместе с тем курица способна к значительным обобщениям, опознавая, например, геометрические фигуры независимо от их цвета. Более того, будучи обучена выбирать сплошной треугольник, она также реагирует на него, когда он предъявляется ей в виде штриховки или даже лишь намечен тремя точками! Так же как и рыбы, птицы способны распознавать относительную величину фигур и так же ошибаются на основе зрительных иллюзий.
Способность к зрительному обобщению у многих позвоночных экспериментально доказана. Особенно в отношении внешних их представителей было показано, что они в состоянии даже в очень сложных ситуациях вычленять существенные детали в воспринимаемых объектах и узнавать эти объекты в сильно измененном виде. Отсюда напрашивается вывод о существовании у позвоночных достаточно сложных общих представлений. Представления чрезвычайно важны для выживания. В них закрепляется в общем виде индивидуальный опыт, что позволяет животному легче ориентироваться в существенно меняющейся окружающей среде при временном отсутствии жизненно важных раздражителей. Подобные зрительные обобщения обнаруживаются, в частности, в явлениях «экстраполяции», описанных Л. В. Крушинским, которые проявляются в том, что в ряде случаев животные способны, проследив за движением какого-либо предмета, ориентироваться затем на ожидаемый путь его перемещения. Следовательно, учитывается предстоящее передвижение объекта после его исчезновения. В проведенных им опытах птицы и млекопитающие помещались перед туннелем, имевшим посредине разрыв, через который можно было наблюдать за движением перемещающейся в туннеле кормушки с приманкой. В других опытах применялась ширма со щелью. Некоторые из подопытных животных (врановые, хищные) обегали затем туннель или ширму по направлению движения приманки и ожидали ее в месте появления. На такое «предвосхищающее» поведение оказались, однако, способными лишь животные, имеющие в естественных условиях дело с прячущейся добычей, которую приходится выслеживать или преследовать, зачастую обгоняя ее, или же сами прячущие корм про запас. Как уже раньше указывалось, мы имеем здесь дело с широко распространенной среди высших позвоночных способностью к нахождению обходного пути. При этом важно отметить, что в любом случае, когда животное огибает непрозрачную преграду или даже просто отворачивается от целевого объекта, оно на время теряет последний из виду. Более того, каждое представление по самой природе своей требует отсутствия ранее воспринятых объектов.
Убедительные доказательства тому, что обобщенные зрительные образы в форме представлений широко распространены среди позвоночных, дали эксперименты, проведенные методом отсроченных реакций (исследования В. С. Хантера, Р. Йеркса, О. Л. Тинклпоу, Ф. Бойтендайка, В. Фишеля и др.). В Советском Союзе такие эксперименты проводились Н. Ю. Войтонисом, И. С. Беритовым, Г. З. Рогинским и др. Обычная схема проведения подобных опытов такова: на виду у животного прячется корм, который, однако, позволяется ему отыскать лишь некоторое время спустя. Обычно подопытное животное должно произвести выбор между несколькими местами, где может находиться спрятанная приманка. При успешном решении задачи имеет место реакция на отсутствующий стимул, что возможно лишь при наличии чувственных представлений.
Опыты по методу отсроченных реакций с успехом ставились над различными позвоночными, особенно же часто над обезьянами. Весьма эффектными были опыты, которые проводил Тинклпоу, когда на глазах у шимпанзе прятались фрукты (бананы), которые затем незаметно для животного подменялись значительно менее привлекательным кормом — салатом или капустой. Найдя спустя положенное время зелень; обезьяна медлила брать ее и продолжала, иногда с визгом, поиски. Последние продолжались до 33 секунд, в то время как в контрольном опыте, когда обезьяне с самого начала показывался салат, который затем не подменялся, поиски продолжались лишь 3 секунды и обезьяна спокойно съедала его. Напрашивается вывод, что обезьяна ищет виденный ею вначале объект, руководствуясь зрительным представлением, которое, по Тинклпоу, отражает также определенные количественные и качественные показатели.
Войтонис на основании своих экспериментов также пришел к выводу, что обезьяны (макаки и шимпанзе) реагируют не на кормушку, содержащую приманку, а на корм как таковой, точнее, на специфический вид корма. То же самое относится и к собакам, но не к рыбам (опыты Бойтендайка и Фишеля).
Наличие у позвоночных (а возможно, и у некоторых высших беспозвоночных) представлений, выражающихся в отсроченных реакциях и способности к нахождению обходных путей (включая и явления экстраполяции), придает их поведению исключительную гибкость и намного повышает эффективность их действий на поисковых этапах поведенческих актов. Вместе с тем не следует переоценивать эти способности, ибо они не обязательно свидетельствуют о высоком уровне психического развития. Основываясь на развитой памяти, они встречаются у позвоночных на разных филогенетических ступенях в зависимости от образа жизни животного: наряду с животными, прячущими в разных местах запасы, эти способности свойственны хищникам, охотящимся на труднодосягаемых, прячущихся или лишь эпизодически появляющихся на виду животных.
На эту биологическую обусловленность реакции в отсутствии стимула, который до этого ее обусловливал, указал еще полвека тому назад советский зоолог и зоопсихолог Д. Н. Кашкаровна примере кошки, которая «стережет мышь и видит, что мышь появляется из норы. Мышь исчезает прежде, чем кошка начнет реагировать. Однако кошка встает и идет к норе. Здесь не может быть вопроса о том, что в момент реакции определяющий стимул отсутствует… Правильный выбор между тремя норами кошка могла бы сделать лишь на основании непосредственно предыдущего появления мыши, т. е. на основе представления о последней».[55] Что же касается обезьян, то, как видно из упомянутых исследований, мы явно имеем здесь дело с более сложными комплексными процессами.
Способность высших позвоночных к предметному восприятию, способности к анализу и обобщению, а тем самым к формированию представлений являются важной предпосылкой образования сложных навыков, которые уже рассматривались раньше (см. ч. I) и которые составляют основное содержание накопления индивидуального опыта не только в сенсорной, но и в эффекторной сфере, точнее сказать, в моторно-сенсорной сфере, действующей как единая система.
Общение
У высших позвоночных особой сложности достигают и процессы общения. Как и у других животных, средства коммуникации включают у них элементы различной модальности — ольфакторные, тактильные.
Ольфакторная сигнализация, т. е. передача информации другим особям химическим путем, преобладает в территориальном поведении, особенно при маркировке местности у хищных и копытных. Для этого служат специальные железы, расположенные в различных участках кожи и выделяющие специфический пахучий секрет. Последний распространяется по воздуху или наносится на различные предметы (стволы и ветки растений, камни и т. д.). Пахучие метки производятся также с помощью испражнений. Выделениями пахучих желез маркируется и след, что способствует нахождению друг друга особями одного вида. Однако во всех этих случаях общение существенно не отличается от того, что мы наблюдаем, скажем, у насекомых.
Видотипичные, инстинктивные компоненты поведения позвоночных, служащие для акустического и оптического общения между животными, как правило, ритуализованы. Оптическое общение осуществляется прежде всего с помощью выразительных поз и телодвижений, о чем уже говорилось выше. Видоспецифические отличия выражаются часто в малозаметных человеку, но четко генетически фиксированных деталях, вызывающих совершенно специфические, зачастую ритуализованные реакции у особей того же вида. Особенно часто такие различия наблюдаются у близкородственных животных, причем прежде всего в сигнальных системах, связанных с размножением. Здесь они служат одним из факторов биологического разграничения видов.
Конкретные формы оптического общения отличаются у высших позвоночных большим разнообразием и дифференцированностью. Нередко они выражаются в специфических «диалогах» между двумя или несколькими особями, т. е. во взаимной демонстрации частей тела или поз. Особенно это относится к «ритуалам» запугивания или «импонирования». «Импонирование» выражает превосходство одной особи по отношению к другой, имеет и оттенок угрозы и вызова, но, в сущности, не носит агрессивного характера (в отличие от подлинного «запугивания», за которым часто следует нападение). Часто самцы «импонируют» самкам во время «ухаживания» за ними. Подчиненная особь отвечает на «импонирование» позой «покорности», чем окончательно исключается возможность столкновений между животными.
У млекопитающих оптическое общение часто сочетается с ольфакторным (рис. 43), так что выделение систем общения по отдельным модальностям является у этих животных в большой степени условным.
Это относится и к акустическому общению, в качестве примера которого следует, конечно, прежде всего напомнить о криках и песнях птиц. Хотя человеческое ухо неспособно улавливать все издаваемые птицами звуки (значительная часть их находится в ультразвуковом спектре ниже 50 кГц), мы поражаемся огромному разнообразию их голосовых реакций. К последним необходимо добавить и неголосовые звуки, как, например, свист крыльев при полете, постукивание дятла по стволу деревьев и т. д. Видоспецифичность таких звуков делает их пригодными для общения.

Для биологически адекватного реагирования на все эти звуковые сигналы существенной является видоспецифическая настройка слуховой системы на определенную часть спектра. Как показал советский исследователь биоакустики птиц В. Д. Ильичев, важнейшую роль играют в этой настройке периферические отделы слухового анализатора, выполняющие роль видотипических биологических фильтров. С их помощью осуществляется и специальная «подстройка» слуховой системы на особенно важные для особи звуки. Таким образом, здесь четко выступает характерная для всего инстинктивного поведения взаимообусловленность и единство врожденных функциональных (физиологических и этологических) и морфологических элементов.
Зоосемантика звуковых сигналов птиц охватывает практически все сферы их жизнедеятельности. Эти сигналы служат опознаванию особи (в частности, ее видовой принадлежности), оповещению других особей о физиологическом состоянии экспедиента (сюда относится, например, «попрошайничество») или об изменениях в окружающей среде (появление врага или, наоборот, кормового объекта и т. п.). При этом богатые зоопрагматические средства птицы позволяют экспедиенту передавать перцепиенту весьма детализованную информацию. Так, грач, подлетающий с кормом к гнезду, оповещает об этом насиживающую самку криком, могущим иметь четыре разных оттенка, на которые самка также отвечает разными звуками. Эти оттенки голосовых реакций птиц подчас настолько тонки, что не воспринимаются человеческим ухом.
Очень большое значение имеют не только четкие межвидовые различия акустического общения, но и индивидуальные, по которым особи узнают друг друга (самец — самку, птенцы — родителей и друг друга, члены стаи — друг друга и т. п.). Интересно, например, что, по данным В. Г. Торпа, в ряде случаев совместно поющие партнеры откликаются только на голос друг друга, но не реагируют на пение других особей. Особенно индивидуально различными являются территориальные крики птиц, оповещающих о занятости участка.
Большого разнообразия звуки достигают и у таких птиц, как куриные. Это обусловлено жизнью диких кур в труднопросматриваемых густых кустарниковых зарослях, где условия для оптического общения особенно неблагоприятны. Наиболее громкие и «впечатляющие» звуки куриных птиц — это крики петухов. Их слышимость (для человеческого уха) достигает 2 километров. В отличие, например, от звуков, с помощью которых петух подзывает кур к найденному им корму, кукареканье производится однократно, но зато дольше и на более высоких частотах. Это относится и к звуковым сигналам тревоги (например, при обнаружении врага), испуга или угрозы. Чем выше тональность, громкость и длительность этих сигналов, тем сильнее их эффект. Свое «кукареку» петух кричит на самых «высоких нотах», но всегда одинаковым образом: отклонения не превышают полтона.
Кукареканье — это своего рода вызов другим петухам, и те отвечают ответными криками, правда, лишь в том случае, если соперники — петухи одного «ранга». Если же в пределах видимости закричит петух, занимающий подчиненное положение, то господствующий попросту атакует его, «не удостаивая» ответным криком. Когда же перекликаются петухи, разделенные большим расстоянием и не видящие друг друга, то это типичные территориальные крики, т. е. акустическая маркировка местности, оповещение о занятости участков стайками кур — петухом и его «гаремом». Большое же биологическое значение крика заключается в том, что он предотвращает или хотя бы уменьшает частоту петушиных боев, а вместе с тем обеспечивает захват, освоение и защиту участков, а в конечном итоге расселение вида, но безвредным путем, ибо дело ограничивается одним запугиванием. Петух-соперник заранее предупреждается о том, что данный участок уже занят и ему придется искать другое пригодное для поселения место. И только в том случае, если все такие места уже заняты, начнутся бои за овладение участками. В этом случае победитель — животное более сильное не только в физическом, но и в психическом отношении — станет продолжателем рода, и это, конечно, тоже выгодно для процветания вида, для его прогрессивной эволюции. Аналогичную роль играет у других птиц пение. У домашних же кур крестьянский двор и прилегающие места остаются для петуха его участком, о занятии которого он непрестанно напоминает криком, хотя это поведение уже потеряло свое биологическое значение, поскольку человек обеспечивает все потребности домашних животных и управляет эволюцией вида. Надо думать, что чрезвычайная близость расположения участков домашних петухов заставляет их интенсивнее кукарекать, чем это имело бы место в дикой природе, так как в селе популяция кур «сверхплотная».
Мы не останавливаемся на звуковом общении млекопитающих, так как у них господствуют те же закономерности, что и у птиц.
Ригидность и пластичность в поведении высших позвоночных
Как уже указывалось, вопреки еще распространенному мнению инстинктивное поведение не теряет своей значимости в процессе эволюции, так как оно принципиально не может замещаться научением. Подчеркнем еще раз, что инстинктивное поведение является видовым поведением, научение — индивидуальным, и поэтому ничем не оправдано противопоставление этих двух основных категорий психической деятельности в качестве якобы различных генетических ступеней. На самом деле, как мы могли убедиться, прогресс инстинктивного поведения, в частности, в ряду позвоночных неразрывно связан с прогрессом индивидуально-изменчивого поведения, поэтому позвоночные с высокоразвитыми формами научения обладают и не менее развитыми сложными формами инстинктивного поведения.
Вместе с тем темпы эволюционных преобразований и специфическая роль в эволюции у инстинктивного поведения и научения различны. Главным образом, как показал Северцов, это относится к высшим позвоночным, особенно млекопитающим, у которых психика приобретает значение решающего фактора эволюции благодаря сильному развитию процессов научения, в том числе в высших их проявлениях — интеллектуальных действиях. Но при этом сохраняется в полной мере значение инстинктивной основы поведения, равно как медленно совершающихся изменений инстинктивного поведения. Приведенные примеры в достаточной мере показывают большое разнообразие истинных инстинктивных движений у высших позвоночных, вполне видотипичных, генетически фиксированных и достигающих в ряде случаев большой сложности. Конечно, и у высших позвоночных инстинктивные компоненты служат прежде всего для пространственно-временной ориентации наиболее жизненно важных поведенческих актов. Пространственная ориентация осуществляется и здесь на основе таксисов: тропо-, тело- и менотаксисов, т. е. типичных врожденных элементов поведения, к которым, однако, прибавляются мнемотаксисы, характеризующиеся индивидуальным запоминанием ориентиров. В последнем случае мы уже имеем дело с накоплением индивидуального опыта, и поскольку именно мнемотаксисы играют в жизни высших позвоночных особенно большую роль, то, следовательно, врожденные способы ориентации существенно обогащаются здесь индивидуально приобретаемыми способами.
В этой связи следует отметить, что, как и у других животных, биологическая адекватность реагирования на компоненты окружающей среды обеспечивается и у высших позвоночных именно инстинктивными элементами поведения. Биологически значимые объекты встречаются в окружающей животное среде в весьма разнообразных и, главное, постоянно меняющихся видах. Мы уже знаем, что их потребление или избегание, т. е. адекватное реагирование на биологические ситуации, возможно лишь в том случае, если животное руководствуется некими постоянными признаками этих объектов и ситуации. Именно это и происходит на генетически фиксированной, врожденной основе, когда животное реагирует на ключевые раздражители. К высшим позвоночным это относится точно так же, как к нижестоящим животным. Но у первых реакции на ключевые раздражители в очень большой степени конкретизируются индивидуальным опытом, дополнительно ориентируются хорошо развитым предметным восприятием. Тем самым инстинктивные действия приобретают и известное познавательное значение для животного, ибо помогают ему при ознакомлении с окружающей действительностью.
Особенно высокого уровня развития инстинктивное поведение достигает у высших позвоночных в ритуализованном общении животных друг с другом. Именно в сфере общения инстинктивные формы поведения достигают наибольшей стереотипности. Совершенно ясно, что иначе, без жестко фиксированных зоопрагматических средств, не может быть взаимопонимания между животными, т. е. не может быть подлинной передачи информации. Вместе с тем именно полноценное общение является необходимым условием для высшей интеграции в области поведения — интеграции поведения отдельных особей и целых сообществ.
Однако даже в сфере общения наличие и важное значение благоприобретаемых компонентов не вызывает сомнения. Так, у многих птиц птенцы не смогут петь видотипичным образом, если своевременно не слышали пения своих родителей. Несомненна роль научения и в образовании индивидуальных особенностей звукового общения, а также в многочисленных случаях акустического подражания у взрослых птиц. В последнем случае оказалось, что заимствованные звуки могут служить для общения наряду с собственными. На этой основе развивается межвидовое общение птиц, т. е. передача информации между особями разных видов. Здесь индивидуально приобретенные коммуникативные компоненты с четким сигнальным значением уже не только модифицируют и обогащают собственно инстинктивные компоненты, но и выступают во вполне самостоятельной роли.
Индивидуально приобретаемые элементы общения представляют особый интерес, так как показывают возможность выхода за пределы закрытых коммуникативных систем животных. Особенно это относится к общению животных с человеком, в частности при дрессировке служебных собак и т. п. Вероятно, расширенное общение с животными сыграло в свое время немаловажную роль и при одомашнивании диких видов. Все это, разумеется, оказалось возможным лишь на высшем уровне перцептивной психики, т. е. у высших позвоночных. Если же иметь в виду предысторию антропогенеза, о чем пойдет речь ниже, то нельзя не признать, что способность высших позвоночных к расширению своих коммуникативных возможностей путем научения должна была стать важной предпосылкой зарождения человеческих форм общения.
Сказанное здесь о роли благоприобретаемых компонентов в инстинктивном поведении высших позвоночных вполне относится и к другим сферам поведения. Напомним, например, что у зубров и бизонов видотипичные признаки поведения появляются лишь на определенном этапе онтогенеза. Хотя здесь, несомненно, имеет место созревание некоторых видотипичных двигательных компонентов вне зависимости от частных внешних условий, все же и в этих процессах участвуют элементы научения. В остальном ограничимся здесь отсылкой к тому, что раньше говорилось по этому поводу об онтогенезе поведения.
Нет также надобности еще раз говорить о том, что в разных поведенческих актах удельный вес врожденных и благоприобретаемых компонентов различен, что и в навыках позвоночных содержатся качественно-гетерогенные элементы. Напомним лишь, что, не говоря уже о том, что навыки формируются на основе безусловнорефлекторных процессов, в их состав всегда входят консервативные двигательные компоненты, придающие навыкам в некоторых отношениях сходство с инстинктивными действиями. Более того, именно формирование таких консервативных компонентов составляет во многих случаях конечный итог и биологическую сущность образования навыка. Это. относится к двигательным стереотипам, автоматизмам, возникающим как результат закрепления навыка в ходе тренировки. Столь примитивные навыки встречаются не только у низших позвоночных. Наоборот, заученные автоматизированные действия играют немаловажную роль и в жизни высших млекопитающих, включая обезьян, а также человека (например, техника письма, повседневные «бытовые» движения и т. п.). У рыб же такими элементарными навыками, очевидно, ограничиваются их способности к научению. С этим связан и тот факт, что у рыб крайне трудно, если вообще возможно переделать положительное значение раздражителя на отрицательное и наоборот. Причиной тому является, в частности, недостаточное развитие ассоциативных функций головного мозга низших позвоночных. Как показал советский физиолог Б. Ф. Сергеев, только на уровне костистых рыб и земноводных появляется способность к образованию временных связей между всеми анализаторными системами и функциями организма, но еще отсутствуют внутри- и межанализаторные ассоциативные временные связи. Анализаторные системы еще разобщены, локализованы в различных отделах мозга.
Только у птиц и млекопитающих замыкательная функция головного мозга получает свое полное развитие, поэтому сложные навыки, в которых решающее значение имеют лабильные, гибкие компоненты, встречаются только у высших позвоночных. Они и определяют далеко идущую изменчивость, пластичность всего поведения животного, чему придавал столь большое адаптационное значение А. Н. Северцов. И действительно, сложные пластичные навыки в полной мере выполняют функцию быстрого приспособления организма к быстротечным изменениям среды. Пластичность навыков высшего порядка дополняет ригидность навыков низшего порядка и инстинктивных действий.
Эта пластичность проявляется в возможности быстрой перестройки навыка, в частности при превращении положительного или отрицательного раздражителя в противоположный. Другая важная особенность — возможность переноса навыка в новые условия, другими словами, адекватное использование накопленного чувственного и моторного опыта при существенных изменениях условий среды. Обеспечиваются эти возможности сильным развитием пластичности в сенсорной сфере, способностью к широким чувственным обобщениям, о чем уже шла речь выше.
Поясним сказанное на нескольких примерах. В первом эксперименте крыса получает приманку в результате ряда последовательных сложных инструментальных действий: она взбирается по лестнице на площадку, поднимает там эту лестницу с помощью перекинутой через блок бечевки, затем поднимается по лестнице на следующую площадку и там получает пищевое подкрепление. К этим фазам можно было бы добавить еще несколько — суть дела от этого не меняется: мы имеем здесь сложную операцию, в которой последовательность отдельных действий животного предопределена и строго фиксирована условиями задачи, т. е. теми компонентами среды (преградами), на которые по необходимости направлена активность животного.
В другом примере при относительно простой структуре операции выделяется способность к далеко идущему переносу операции в измененных условиях, что также весьма характерно для сложных навыков. В данном случае крыса, научившаяся находить кратчайший путь к «цели» в лабиринте, оказалась без какого-либо нового обучения способной к этому и после того, как лабиринт был поставлен вертикально. Поскольку животное при этом руководствуется и новыми ощущениями (с вестибулярного аппарата), то здесь с очевидностью имеют место и явления переноса в сенсорной сфере.
В третьем эксперименте крыса, освоившая сухопутный лабиринт, тут же с неменьшим успехом проплыла этот лабиринт после того, как он был заполнен водой. Поскольку при плавании производятся иные движения, чем при ходьбе, мы можем здесь констатировать далеко идущий перенос в моторной сфере при сохранении прежней ориентации в пространстве.
Даже в первом примере, а тем более в остальных, невозможно расценивать поведение подопытного животного как простую цепь механически усвоенных движений, ибо в таком случае совершенно исключалась бы возможность переноса операции и сохранения навыка в измененных условиях. Сложные навыки представляют собой исключительно динамические моторнорецепторные системы, обеспечивающие на основе высокоразвитой ориентировочной деятельности выработку весьма пластичных двигательных программ. Процесс ориентировки сливается здесь с двигательной активностью, а нахождение верного решения задачи формируется в ходе этой активности на основе высокоразвитого чувственного обобщения.
Отмеченными качествами сложные навыки выделяются среди других видов научения, и именно благодаря им сложные навыки стали предпосылками и основой развития высших форм психической деятельности животных — интеллектуальных действий.
Проблема интеллекта животных
Предпосылки и элементы интеллектуального поведения животных
Интеллектуальное поведение является вершиной психического развития животных. Однако, говоря об интеллекте, «уме» животных, их мышлении необходимо прежде всего отметить, что чрезвычайно трудно точно указать, по поводу каких животных можно говорить об интеллектуальном поведении, а по поводу каких — нет. Очевидно, речь может идти лишь о высших позвоночных, но явно не только о приматах, как это до недавнего времени принималось. Вместе с тем интеллектуальное поведение животных является не чем-то обособленным, из ряда вон выходящим, а лишь одним из проявлений единой психической деятельности с ее врожденными и благоприобретаемыми аспектами. Интеллектуальное поведение не только теснейшим образом связано с разными формами инстинктивного поведения и научения, но и само складывается (на врожденной основе) из индивидуально-изменчивых компонентов поведения. Оно является высшим итогом и проявлением индивидуального накопления опыта, особой категорией научения с присущими ей качественными особенностями. Поэтому интеллектуальное поведение дает наибольший приспособительный эффект, на что и обратил особое внимание А. Н. Северцов, показав решающее значение высших психических способностей для выживания особей и продолжения рода при резких, быстро протекающих изменениях в среде обитания.
Предпосылкой и основой развития интеллекта животных — во всяком случае в направлении, ведущем к человеческому сознанию — является манипулирование, причем прежде всего с биологически «нейтральными» объектами. Особенно, как уже было показано, это относится к обезьянам, для которых манипулирование служит источником наиболее полных сведений о свойствах и структуре предметных компонентов среды, ибо в ходе манипулирования происходит наиболее глубокое и всестороннее ознакомление с новыми предметами или новыми свойствами уже знакомых животному объектов. В ходе манипулирования, особенно при выполнении сложных манипуляций, происходит обобщение опыта деятельности животного, формируются обобщенные знания о предметных компонентах окружающей среды, и именно этот обобщенный двигательно-сенсорный опыт составляет главнейшую основу интеллекта обезьян.
О манипулировании человекообразных обезьян «биологически индифферентными» предметами Павлов говорил: «Это же — самая настойчивая любознательность. Так что нелепое утверждение, будто у животных ее нет, нет в зачатке того, что есть у нас и что в конечном счете создало науку, — не отвечает действительности».[56] В качестве примера Павлов сослался на наблюдавшееся им у шимпанзе манипулирование предметами, в частности коробкой, в которой нет «никаких апельсинов, ни яблок». Тем не менее обезьяна «долгое время возится… над решением механических задач, которое не обещает ей никаких выгод, никакого материального удовлетворения».[57]
Эта, по Павлову, «чистейшая, бескорыстная любознательность» и заставляет обезьяну изучать объект манипулирования в ходе активного воздействия на него. При этом одновременно и во взаимодействии друг с другом включаются в познавательную деятельность животного разные сенсорные и эффекторные системы. Ведь манипулирующая обезьяна почти непрерывно следит за движениями своих рук; под пристальным зрительным контролем производятся самые разнообразные действия как без разрушения целостности объекта: поворачивание в разные стороны, облизывание, поглаживание, придавливание, перекатывание и т. п., так и деструктивного порядка: разламывание, разрывание, вычленение отдельных деталей и т. д. (рис. 24).
Особую познавательную ценность представляют деструктивные действия, так как они позволяют получить сведения о внутренней структуре предметов. При манипулировании животное получает информацию одновременно по ряду сенсорных каналов, но преобладающее значение имеет у обезьян сочетание кожно-мышечной чувствительности рук со зрительными ощущениями. Кроме того, в обследовании объекта манипулирования участвуют также обоняние, вкус, тактильная чувствительность околоротовых вибрисс, иногда слух и т. д. Эти виды чувствительности сочетаются с кожно-мышечной чувствительностью эффекторов (ротового аппарата, передних конечностей) кроме обезьян и у других млекопитающих, когда те манипулируют предметами. В итоге животные получают комплексную информацию об объекте как едином целом и обладающем разнокачественньми свойствами. Именно в этом и заключается значение манипулирования как основы интеллектуального поведения.
Необходимо, однако, подчеркнуть, что первостепенное значение имеют для интеллектуального поведения зрительные восприятия и особенно зрительные обобщения, о которых уже раньше шла речь. Насколько развита способность к формированию обобщенных зрительных образов даже у крыс — показывает следующий эксперимент, в котором крысы с успехом решали очень трудную задачу: животному надо выбрать из трех предъявленных фигур (вертикальные и горизонтальные полосы) одну несходную по сравнению с двумя другими. Местонахождение и рисунок такой фигуры постоянно меняются, следовательно, это будут то вертикальные, то горизонтальные полосы, расположенные то слева, то справа, то посередине (в неправильной последовательности). Таким образом, подопытное животное могло ориентироваться лишь по одному, крайне обобщенному признаку — несхожести одного рисунка по сравнению с остальными. Мы имеем здесь, таким образом, дело со зрительным обобщением, близким к абстрагированию, свойственному мыслительным процессам.
С другим элементом интеллектуального поведения, на этот раз в двигательной сфере, мы встречались при описании опытов с «проблемными ящиками». И здесь мы имеем дело со сложными многофазовыми навыками, поскольку у высших млекопитающих, например енотов, можно сравнительно легко добиться решения задач, при которых животному приходится в определенной последовательности открывать совокупность разнообразных запирающих приспособлений. Как и при описанном опыте с подтягиванием крысой лестницы, енот может решить такую задачу лишь при соблюдении строго определенной последовательности действий. Но разница заключается в том, что в отличие от крысы еноту приходится самому находить эту последовательность, и это поднимает его деятельность, безусловно, на более высокий уровень. Правда, по некоторым данным и крысы способны на это.
Необходимо, однако, подчеркнуть, что даже высшие позвоночные решают инструментальные задачи труднее, чем локомоторные. Л. Кардош отметил в этой связи, что в психической деятельности животных преобладает познавание пространственных отношений (см. ч. I, гл. 3), постигаемых ими с помощью локомоторных действий. У обезьян же, особенно человекообразных, локомоторное познавание пространственных отношений теряет свою доминирующую роль за счет сильного развития манипуляционных действий. Однако только человек может полностью освободиться от направляющего воздействия пространственных отношений, если этого требует познание временно-причинных связей.
Решение многофазовых инструментальных задач у обезьян изучалось рядом исследователей, в частности Н. Н. Ладыгиной-Котс. В своей монографии «Приспособительные моторные навыки макака в условиях эксперимента» она подвела итог многочисленным опытам, в которых использовались весьма разнообразные сочетания запирающих механизмов. Эти эксперименты показали, что низшая обезьяна (макак-резус) способна научиться отмыкать большие серии запирающих механизмов, хотя лучше справляется с одиночными установками. Характерными явились при этом многочисленные и многообразные нащупывающие движения рук, «экспериментирование». Ввиду ее большой торопливости наиболее трудными оказались для обезьяны те приспособления, которые не отмыкались легкими быстрыми движениями. Легче всего производились такие движения, как вытягивание, отведение, притягивание, опускание и др., труднее всего — отодвигание и вращение, вывертывание. Вообще в поисках пунктов задержки и пути преодоления препятствий преобладающая роль принадлежала кинестетическим, а не зрительным восприятиям. Интересно, что по многим из этих особенностей, как показали более поздние исследования, действия низших обезьян в опытах с запирающими механизмами напоминают таковые енотов.
Чрезвычайно важной предпосылкой интеллектуального поведения является и способность к широкому переносу навыков в новые ситуации. Эта способность вполне развита у высших позвоночных, хотя и проявляется у разных животных в разной степени. В. П. Протопопов приводит следующий пример переноса приобретенного опыта в новую ситуацию у собаки. Первоначально подопытная собака научилась открывать нажатием лапы щеколду на дверце «проблемной клетки», в которой находилась приманка. В других опытах та же собака научилась затем подтягивать зубами и лапами кусок мяса за веревку, которая лежала перед ней на полу. После этого была создана третья ситуация, содержавшая элементы первых двух: на клетке, применявшейся в первой ситуации, щеколда была поднята на такую высоту, что собака не могла ее достать лапой, но к щеколде привязывалась веревка, потянув за которую можно было ее открыть. Когда собака была подведена к клетке, она сразу же, без всяких других проб, схватила зубами веревку и, потянув, открыла щеколду. Таким образом, задача была немедленно решена в новой ситуации, несмотря на то что прежние элементы располагались в ней совсем по-иному: веревка висела, а не лежала горизонтально на полу, на конце ее было привязано не мясо, а щеколда, которая к тому же находилась на другом месте — наверху. К тому же щеколда отпиралась в первых опытах движением лапы, а затем — с помощью зубов. «…Новый навык, — пишет по этому поводу Протопопов, — вырабатывается сразу, „внезапно“, но эта внезапность… обусловлена вполне определенными следами прошлого опыта, которые под влиянием стимула вступают путем как бы взрывчатого замыкания в новую временную связь, и создается новая нервная структура и новая реакция, отличающаяся от прежних двух и в рецепторной и эффекторной части… Подобные навыки… могут по своему внешнему проявлению имитировать разумное поведение и, если не знать этапов их возникновения, можно прийти к ошибочным антропоморфическим заключениям».[58]
К подобным антропоморфическим выводам пришел, например, Н. Р. Ф. Майер, признавший крыс способными «рассуждать». Обоснованием этого заключения послужили результаты его экспериментов по выработке у крыс отсроченных реакций, в ходе которых эти животные оказались в состоянии связывать элементы прежнего опыта, которые раньше никогда не сочетались в их поведении. Как было показано, это происходило и в опытах Протопопова с собакой.
Итак, способности высших позвоночных к разнообразному манипулированию, к широкому чувственному (зрительному) обобщению, к решению сложных задач и переносу сложных навыков в новые ситуации, к полноценной ориентации и адекватному реагированию в новой обстановке на основе прежнего опыта являются важнейшими элементами интеллекта животных. И все же сами по себе эти качества еще недостаточны, чтобы служить критериями интеллекта, мышления животных. Тем более, как указывалось, невозможно признать такими критериями, например, высокоразвитые способности к оптическому обобщению у пчел.
Критерий интеллектуального поведения животных
Отличительная особенность интеллекта животных заключаются в том, что в дополнение к отражению отдельных вещей возникает отражение их отношений и связей (ситуаций). Отчасти это имеет, конечно, место и при некоторых сложных навыках, что лишний раз характеризует последние как переходную форму к интеллектуальному поведению животных. Это отражение происходит в процессе деятельности, которая по своей структуре, согласно Леонтьеву, является двухфазной.
Мы уже видели, что сложные навыки животных большей частью являются многофазными. Однако эти фазы, будь это вскарабкивание крысы с площадки на площадку с помощью подтягиваемой лестницы или последовательное отмыкание затворов «проблемного ящика», по существу своему являются лишь цепью, суммой однозначных равнокачественных этапов последовательного решения задачи. По мере же развития интеллектуальных форм поведения фазы решения задачи приобретают четкую разнокачественность: прежде слитая в единый процесс деятельность дифференцируется на фазу подготовления и фазу осуществления. Именно фаза подготовления составляет характерную черту интеллектуального поведения. Как указывает Леонтьев, интеллект возникает впервые там, где возникает процесс подготовки возможности осуществить ту или иную операцию или навык.
В конкретных экспериментальных исследованиях двухфазность интеллектуальных действий проявляется, например, в том, что обезьяна достает сперва палку, чтобы затем с помощью этой палки сбить высоко подвешенный плод, как это имело место в широко известных опытах немецкого психолога В. Келера. В других экспериментах обезьяна могла овладеть приманкой лишь в том случае, если сперва оттолкнет ее от себя палкой к такому месту, где ее (после обходного движения) можно достать рукой (рис. 44).
Производилось и много других экспериментов, в которых обезьяны должны были решить задачу с употреблением орудия (чаще всего палки). Так, в опытах Г. З. Рогинского шимпанзе, имевшие опыт манипулирования палками, сразу употребляли таковые для доставания приманки. Но низшие обезьяны, кроме одной (павиан чакма), к этому оказались не сразу способны. Все же Рогинский отвергает мнение В. Келера о наличии разрыва между психикой человекообразных и низших обезьян.
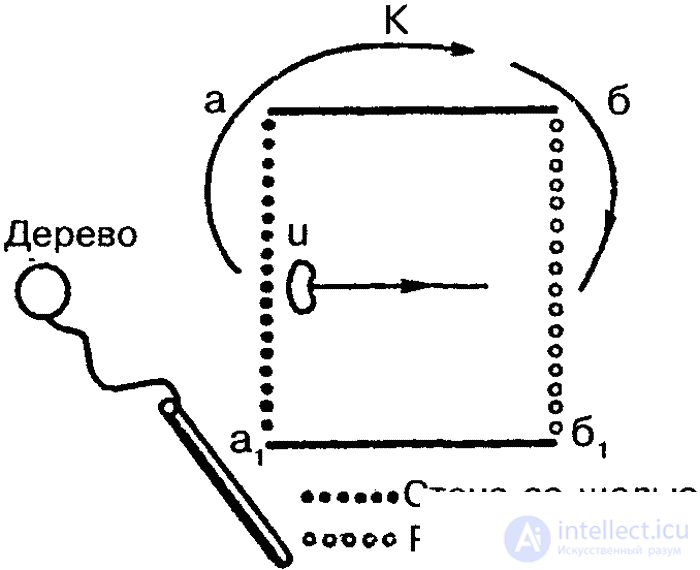
Советский зоопсихолог Л. С. Новоселова сумела своими исследованиями выявить генезис употребления палок при решении сложных задач у шимпанзе. Она показала, что употребление палки формируется как индивидуально-приспособительное действие, но не является врожденной формой поведения. При этом намечается несколько этапов — от оперирования всей рукой как рычагом к специализированным действиям кистью, которая уже не только удерживает палку, но и направляет ее движения в соответствии со специфическими свойствами орудия.
Н. Н. Ладыгина-Котс детально изучала у шимпанзе процесс подготовки и даже изготовления-орудия, необходимого для решения технически несложной задачи — выталкивания приманки из узкой трубки. На глазах у шимпанзе в трубку закладывалась приманка таким образом, что ее нельзя было достать просто пальцами. Одновременно с трубкой животному давались различные предметы, пригодные для выталкивания прикорма после некоторой их «доработки» (рис. 45). Подопытная обезьяна вполне (хотя и не всегда немедленно) справлялась со всеми этими задачами.

В этих опытах также четко выступает двухфазность интеллектуального действия: приготовление орудия — первая, подготовительная фаза, доставание приманки с помощью орудия — вторая фаза. Первая фаза вне связи со следующей фазой лишена какого бы то ни было биологического смысла. Вторая фаза — фаза осуществления деятельности — в целом направлена на удовлетворение определенной биологической потребности животного (в описанных опытах — пищевой).
По Леонтьеву, первая, подготовительная фаза побуждается не самим предметом (например, палкой), на который она направлена, а объективным отношением палки к приманке. Реакция на это отношение и есть подготовка второй фазы, фазы осуществления, которая направлена на объект («цель»), побуждающий всю деятельность животного. Вторая фаза включает, таким образом, в себя определенную операцию, закрепляемую в виде навыка.
Большое значение как один из критериев интеллектуального поведения имеет и то обстоятельство, что при решении задачи животное пользуется не одним стереотипно выполняемым способом, а пробует разные способы, которые являются результатом ранее накопленного опыта. Следовательно, вместо проб различных движений, как это имеет место при неинтеллектуальных действиях, при интеллектуальном поведении имеют место пробы различных операций, что позволяет решить одну и ту же задачу различными способами. Перенос и, пробы различных операций при решении сложной задачи находят у обезьян свое выражение, в частности, в том, что они практически никогда не пользуются орудиями совершенно одинаковым образом.
Таким образом, при интеллектуальном поведении мы имеем дело с переносом операции, причем этот перенос не требует, чтобы новая задача была непосредственно сходной с прежней. Операция, как отмечает Леонтьев, перестает быть неподвижно связанной с деятельностью, отвечающей определенной задаче. И здесь мы можем проследить преемственность от сложных навыков.
Поскольку интеллектуальное поведение животных характеризуется отражением не просто предметных компонентов среды, а отношений между ними, здесь осуществляется и перенос операции не только по принципу сходства вещей (например, преград), с которыми была связана данная операция, но и по принципу сходства отношений, связей вещей, которым она отвечает.
Формы мышления
На основе многолетних экспериментальных исследований Ладыгина-Котс пришла к выводу, что мышление животных имеет всегда конкретный чувственно-двигательный характер, что это мышление в действии, причем эти действия являются всегда предметно-отнесенными. Такое мышление в действиях, по Ладыгиной-Котс, представляет собой практический анализ и синтез, который при орудийной деятельности осуществляется в ходе непосредственного обращения с объектами, в ходе их обследования, обработки и применения. Но при этом не исключаются, а, наоборот, играют большую роль те обобщенные зрительные представления, о которых шла речь выше.
В соответствии с этим Ладыгина-Котс считает возможным выделить две различные по сложности и глубине формы мышления (правда, только у человекообразных обезьян).
Первая форма характеризуется установлением связей раздражителей (предметов или явлений), непосредственно воспринимаемых животным в ходе его деятельности. Это анализ и синтез в наглядно обозреваемой ситуации. Примером может служить выбор обезьяной предметов, пригодных для употребления в качестве орудия, с учетом их величины, плотности, формы и т. д.
Вторая форма характеризуется установлением связей между непосредственно воспринимаемыми раздражителями и представлениями (зрительными следами). Так, в описанных опытах, в которых шимпанзе приходилось выталкивать лакомство из трубы с помощью орудия, мыслительные операции такого типа проявлялись в изготовлении орудия, например в распутывании клубка проволоки и ее выпрямлении. Особенно убедительным доказательством существования этой высшей формы мышления является опыт, при котором обезьяне вместе с трубой давалась дощечка значительно более широкая, чем диаметр трубы. Шимпанзе оказался в состоянии совершенно самостоятельно отчленять от доски узкие лучины и использовать их в качестве орудия для выталкивания приманки из трубы.
Подобное поведение обезьяны можно здесь объяснить только тем, что у нее в ходе предыдущей деятельности сформировалось обобщенное зрительное представление о предмете типа палки (но только в ситуации данной задачи). Решающая роль предшествующего опыта в формировании подобных зрительных «представлений» достаточно четко выступает в этой же серии экспериментов. Ведь еще до решения описанной задачи обезьяна накапливала опыт по «доработке» весьма различных предметов, среди которых были и фигурные дощечки (рис. 46), превращение которых в пригодные для решения задачи орудия было для шимпанзе делом не более трудным, чем обламывание боковых побегов ветки (рис. 45). Вместе с тем изготовление орудия из такой фигурной дощечки требует отщепления части ее в продольном направлении. Подобные действия и накапливаемый при этом опыт и подготовили обезьяну к решению задачи с широкой дощечкой, которая ей давалась в одном из следующих экспериментов.
На основе установления связи обобщенного зрительного представления о необходимом предмете (типа палки) с непосредственным восприятием второго предмета (трубы), также данного в конкретной ситуации опыта, обезьяна сумела вычленить (по воображаемой линии!) часть из целого — лучину из доски, причем таким образом, что она оказалась пригодной служить орудием для выталкивания приманки.
Таким образом, шимпанзе в состоянии мысленно разлагать целые объекты на детали, равно как и сложные фигуры — на их составные части. Как уже указывалось, ведущую роль в поведении, и в частности в интеллектуальных действиях обезьян играют их руки, тактильно-кинестетическая чувствительность руки. И. П. Павлов говорил поэтому с полным основанием о «ручном мышлении» обезьян. Сочетание тактильно-кинестетической чувствительности со зрением дает обезьяне большие преимущества для установления пространственно-временных связей для практического анализа и синтеза. Именно это существенное расширение и углубление сенсорной сферы легло у обезьян в основу того, что И. П. Павлов обозначил как «улавливание постоянной связи между вещами» (или «нормальной связи вещей»).[59]
Биологическая ограниченность интеллекта животных
Наряду со всем этим надо четко представить себе биологическую ограниченность интеллектуального поведения обезьян. Как и все другие формы поведения, оно всецело определяется образом жизни и чисто биологическими закономерностями, рамки которых не может перешагнуть даже самая умная обезьяна. Так, например, шимпанзе на воле каждый вечер устраивают себе искусно переплетенные спальные гнезда из ветвей и листьев, но, по свидетельству английской исследовательницы поведения обезьян Дж. ван Лавик-Гудолл, они никогда не сооружают навесов и остаются совершенно беззащитными под проливным тропическим дождем.
Очень редко обезьяны пользуются на воле и орудиями. Правда, имеются отдельные наблюдения об орудийных действиях шимпанзе при добывании пищи или нападении. Но, как и другие человекообразные обезьяны, шимпанзе вполне обходятся в повседневной жизни без орудий. С другой стороны, другие позвоночные (каланы, галапагосские дятловые вьюрки и др.) систематически пользуются предметами в качестве орудий. Уже это указывает на то, что орудийные действия сами по себе не обязательно являются критериями высокоразвитой психической деятельности животных.
Биологическую ограниченность интеллекта антропоидов вскрывает и анализ экспериментальных данных. Так, Ладыгина-Котс показала, что зрительные образы, представления человекообразных обезьян являются значительно более слабыми, чем у человека, и всегда связаны с компонентами окружающей обстановки (ситуационная связанность представлений).
Эта ограниченность интеллектуального поведения неоднократно проявлялась в опытах Ладыгиной-Котс, когда шимпанзе допускал «нелепые» ошибки при употреблении предметов, предоставленных ему для выталкивания приманки из трубы. Так, например, он пытался втолкнуть в трубу кусок фанеры вопреки явному несоответствию ее ширины и принимался обгрызать его лишь после ряда таких неудачных попыток. Подчас неадекватность действий обусловливалась преобладанием деструктивного манипулирования (рис. 47). Кроме того, была проведена специальная серия опытов, в которых обезьяне давалась труба, закрытая с одного конца, и крючок. Если приманка, заложенная в такую трубу, была прикреплена к нити, шимпанзе легко вытягивал ее оттуда. Несмотря на это обезьяна не сумела адекватно применить крючок и, более того, чаще всего отламывала загнутую его часть как мешавший элемент. Ладыгина-Котс писала по этому поводу, что «шимпанзе оказался неспособным перейти от шаблонного привычного приема выталкивания приманки прямым, гладким единичным орудием к использованию приема притягивания к себе крюком», и усматривала в этом «недостаточную пластичность психики шимпанзе, ограниченность его мышления».[60]
Шимпанзе, по Ладыгиной-Котс, «не в состоянии схватить сразу существенные особенности в новой ситуации и установить новые связи на основе осмысливания непосредственно воспринимаемых отношений между предметами».[61]
Этот вывод Ладыгиной-Котс подтверждается экспериментами и других исследователей. Так, шимпанзе показал ситуационную связанность своих представлений и неспособность охватить существенное изменение в прежней ситуации в следующем опыте: шимпанзе предлагается с помощью палки подкатывать яблоко к клетке вокруг невысокой стенки. После овладения обезьяной этим навыком убирается часть стенки непосредственно перед клеткой, в результате чего было бы удобнее непосредственно притягивать яблоко палкой. Тем не менее обезьяна продолжает выполнять прежнее сложное, трудное действие, отталкивая яблоко от себя и обводя его вокруг стенки (опыт Э. Г. Вацуро, рис. 48).

Даже самые сложные проявления интеллекта обезьян представляют собой в конце концов не что иное, как применение в новых условиях филогенетически выработанного способа действия. Ведь давно уже подмечалось сходство притягивания приманки с помощью палки с притягиванием плода, растущего на ветке. Войтонис и Ладыгина-Котс указывали на то, что развитая способность обезьян к практическому анализу связана с особенностями их питания; высокоразвитые сенсомоторные функции руки, их сочетание со зрением и как следствие отличительные познавательные способности обезьян Фабри объясняет функциональными особенностями их хватательных способностей (см. гл. 3) и т. д. Эта биологическая обусловленность всей психической деятельности обезьян, включая и антропоидов, и является причиной отмеченной ограниченности их интеллектуальных способностей, причиной их неспособности к установлению мысленной связи между одними лишь представлениями и их комбинированием в образы. Неспособность же мысленно оперировать одними лишь представлениями с неизбежностью приводит к неспособности понимать результаты своих действий, понимать истинные причинно-следственные связи. Это возможно лишь с помощью понятий, которые у обезьян, как и у всех других животных, по указанным причинам всецело отсутствуют.
В заключение приходится признать, что проблема интеллекта животных еще совершенно недостаточно изучена. По существу до сих пор проведены обстоятельные экспериментальные исследования только над обезьянами, преимущественно высшими, в то время как о возможности интеллектуальных действий у других позвоночных еще почти нет доказательных экспериментальных данных. Вместе с тем, как уже говорилось, сомнительно, чтобы интеллект был присущ только приматам.
Если я не полностью рассказал про перцептивная психика? Напиши в комментариях Надеюсь, что теперь ты понял что такое перцептивная психика и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Сравнительная психология и Зоопсихология

Комментарии