Лекция
Привет, мой друг, тебе интересно узнать все про инстинктивное поведение, тогда с вдохновением прочти до конца. Для того чтобы лучше понимать что такое инстинктивное поведение , настоятельно рекомендую прочитать все из категории Сравнительная психология и Зоопсихология.
инстинктивное поведение как основа жизнедеятельности животных
Как было показано, инстинктивное поведение и научение не существуют в реальном поведении сами по себе, а только совместно, переплетаясь друг с другом в единый поведенческий акт. Однако это не означает, что инстинктивное поведение или научение представляет собой лишь условности, искусственно созданные для научного анализа. Условным является здесь лишь их разделение, сами же эти компоненты действительно существуют и функционируют как четко различимые процессы жизнедеятельности со своими специфическими качественными особенностями.
В современных научных исследованиях все чаще избегают употребления понятия «инстинкт» ввиду многообразия и нечеткости его толкования. Понимаемое же как врожденное, наследственно фиксированное, видотипичное «инстинктивное поведение» складывается из инстинктивных действий или актов, которые в свою очередь состоят из отдельных инстинктивных движений (или поз, звуков и т. п.). Четкое разграничение этих терминов необходимо для этологического анализа обсуждаемой категории поведения животных.
Когда мы говорим, что поведение является совокупностью функций внешних, «рабочих» органов животного организма, необходимо различать сами эти функции и их ориентацию во времени и пространстве. И то и другое происходит на инстинктивной основе. Научением же можно только менять ориентацию этих функций. Это означает, что никаким научением нельзя заставить функционировать органы животного иначе, чем это обусловлено их генетически фиксированным строением. Именно морфологические особенности определяют характер функционирования экзосоматических органов, т. е. инстинктивных движений. Нельзя вопреки поговорке научить зайца спички зажигать, так как у него нет соответствующих морфофункциональных предпосылок в строении его конечностей. Но можно научить зайца пользоваться своими конечностями естественным (инстинктивным) образом в нужный момент и в определенном направлении, т. е. ориентировать его инстинктивные движения во времени и пространстве путем научения (в данном случае — дрессировки).
Сказанное не следует понимать в том смысле, что в поведении вообще первичным являются строение, структура, а вторичным — функция, движение. Наоборот, мы говорим о примате движения, функции, имея при этом в виду, что функция определяет форму. Биологическая обусловленность поведения не означает его морфологическую обусловленность; особенно в историческом, филогенетическом плане. В процессе эволюции, несомненно, поведение определяло формирование морфологических признаков, необходимых именно для более успешного выполнения самих поведенческих актов.
Но когда мы говорим о конкретных инстинктивных движениях — результатах эволюционного процесса, то имеем в виду функции именно этих морфологических образований и то, что форма выполнения поведенческих функций обусловливается соответствующими морфологическими структурами. Конкретно это означает, что каждое животное может передвигаться или питаться только так, как это определяется специфическим строением его внешних органов, служащих для выполнения этих функций.
Учитывая все это, можно сказать, что вся жизнедеятельность животного организма, проявляющаяся во внешней активности, зиждется на инстинктивных движениях и других инстинктивных реакциях (термических, электрических, изменения окраски, выделения секретов и т. п.). Они обеспечивают все жизненные функции организма, процессы обмена веществ, а тем самым существование особи и размножение. Вот почему мы говорим о примате инстинктивных движений и по отношению к нервной деятельности, сенсорике, психическому отражению, которые служат у животных лишь для осуществления этих движений, для их ориентации. Поэтому в эволюционном плане развитие психики являлось необходимым следствием (а затем уже — и предпосылкой) повышения уровня обмена веществ и двигательной активности.
Инстинктивное поведение не исчерпывается, однако, самими функциями экзосоматических органов, а включает в себя и механизмы их регулировки и пространственно-временной ориентации. В этом отношении регулировка и ориентация, осуществляемые благоприобретенным путем, на основе научения, служат хотя и важным, но все же лишь дополнением к этим инстинктивным процессам.
Внутренние факторы инстинктивного поведения
Как уже упоминалось, проблема инстинкта и научения непосредственно связана с другой не менее важной проблемой — проблемой внутренних и внешних факторов, мотивации поведения.
Долгое время считали, что инстинктивные действия определяются внутренними, притом загадочными, причинами, в то время как индивидуальное научение зависит от внешних стимулов. В таком виде представления об исключительном или хотя бы преобладающем значении внутренних или внешних факторов встречаются уже у античных мыслителей. При этом мистический, телеологический подход основывался на постулате изначальной целесообразности чисто внутренних, передаваемых от поколения к поколению факторов. Механистический же подход, получивший особое развитие со времен Декарта, признавал лишь внешние факторы движущими силами поведения. В ряде случаев и тот и другой взгляд непримиримо отстаивались вплоть до нашего века.
Что же мы сегодня знаем о мотивации, о движущих силах инстинктивного поведения, а тем самым о поведении вообще? Обратимся сначала к тем внутренним факторам, которые дают первый толчок к любому поведенческому акту, не вдаваясь при этом в детали тех весьма сложных физиологических процессов, которые удалось в этой связи изучить за последние десятилетия.
Внутренняя среда животного организма непрерывно обновляется, но, несмотря на непрекращающиеся процессы ассимиляции и диссимиляции, эта среда остается по своим физиологическим показателям постоянной. Стабильность внутренней среды — совершенно необходимое условие для жизнедеятельности организма. Только при этом условии могут осуществлятся необходимые биохимические и физиологические процессы. Любые, даже незначительные, отклонения от нормы воспринимаются системой интерорецепторов и приводят в действие физиологические механизмы саморегуляции, в результате чего эти нарушения устраняются. Такими механизмами саморегуляции советский физиолог академик П. К. Анохин считал сложные динамические структуры, функционирующие по принципу обратной связи (обратной афферентации) и обозначенные им как функциональные системы (рис. 5).

Итак, постоянство внутренней среды основано на самовосстанавливающейся уравновешенности внутренних процессов организма. Важная особенность этих процессов состоит в том, что они протекают в форме ритмов, которые также строятся на системах саморегулирования. Именно в сдвигах этих ритмов видный советский зоопсихолог В. М. Боровский еще в 30-е годы усматривал первичную мотивацию поведения. Выступая против идеалистического понимания инстинктивного поведения, он показал, что в мотивации этого поведения, т. е. в том, что принято называть побуждениями или влечениями, нет ничего сверхъестественного, оторванного от материального мира. Внутреннее побуждение, подчеркивал он, всегда является сдвигом корреляции физиологических ритмов в организме в сторону установления наиболее выгодной в данных условиях коррелированности ритмов всех физиологических процессов. В этом постоянном восстановлении внутреннего равновесия Боровский усматривал основу жизнеспособности организмов.
Итак, первопричиной и основой мотивации поведения являются более или менее значительные и длительные отклонения от нормального уровня физиологических функций, нарушения внутренних ритмов, обеспечивающих жизнедеятельность организма. Эти сдвиги выражаются в появлении потребностей, на удовлетворение которых и направлено поведение.
Первостепенное значение имеют для внутренней мотивации поведения ритмические процессы, происходящие в центральной нервной системе. Собственная ритмика ее стволовой части у позвоночных и брюшных нервных структур у беспозвоночных обеспечивает прежде всего ориентацию поведения во времени. Сейчас хорошо известны автономные, самовозбуждающиеся колебательные процессы («внутренние», или «биологические, часы»), которые регулируют общую ритмику жизнедеятельности организма. В отношении поведения это означает, что периодические колебания внешней активности животных, начало и окончание ритмично повторяющихся действий определяются ритмом «внутренних часов», синхронизированных с космическим временем. Существенные поправки или изменения вносятся в поведенческие ритмы многообразными биологически важными факторами среды, но общая «канва» инстинктивного поведения определяется самовозбуждающимися колебательными процессами с периодом, равным приблизительно суткам (околосуточный, «циркадный» ритм).
В нормальных условиях этот ритм синхронизирован с изменениями в среде, определяемыми вращением Земли вокруг своей оси в течение суток. Однако и в искусственных условиях полной изоляции животного можно наблюдать обычную смену форм активности в те же сроки, что и в нормальных условиях. Это могут быть, например, изменения, связанные со сменой дня и ночи, хотя животное находится в эксперименте в условиях постоянного равномерного освещения.
Помимо циркадных ритмов в поведении животных проявляются и более краткосрочные ритмы, многократно повторяющиеся в течение дня. Так, немецкий этолог В. Шлейдт установил, что клохтанье индюка вполне закономерно повторяется периодически даже в том случае, когда птица полностью изолирована от внешнего мира и даже лишена слуха. Конечно, в нормальных условиях протекание внутренних ритмов изменяется под влиянием внешних воздействий (слуховых, зрительных и прочих стимулов, метеорологических факторов и т. д.), а также зависит от общего физиологического состояния животного.
«Внутренние часы» необходимы и для ориентации животных в пространстве. Хороший пример тому — ориентация птиц во время перелета. Руководствуясь, например, таким астрономическим ориентиром, как солнце, птицы должны учитывать его положение на небосводе в каждое данное время дня, что и происходит путем сопоставления воспринимаемой информации о положении солнца с фазами циркадного ритма.
Уже упомянутый эксперимент Шлейдта показывает, что эндогенные процессы в нервной системе способны обусловливать выполнение определенных инстинктивных движений и при полном отсутствии адекватных внешних стимулов. Так, немецкий нейрофизиолог Э. Хольст обнаружил в стволовой части головного мозга курицы ряд зон, активация которых (в эксперименте — электрическим током) вызывает типичные инстинктивные движения различного функционального значения. При этом оказалось, что при раздражении одного и того же участка мозга, с усилением силы раздражения одно инстинктивное действие сменяется другим в естественной последовательности. Получилась цепь видотипичных движений, выполняемых курицей в определенной биологически значимой ситуации, например при виде приближающегося к ней наземного врага. При этом не только выполнение двигательных реакций, но и последовательность их появления точно так же соответствовала естественному поведению курицы: сперва лишь легкое беспокойство, потом подъем, усиленное беспокойство и, наконец, взлет (рис. 6). Учитывая, что все это происходило в отсутствие каких-либо адекватных внешних раздражителей, становится ясным, что на сугубо эндогенной основе могут выполняться не только отдельные инстинктивные движения, но и целые системы таких движений — инстинктивные действия. Конечно, в естественных условиях такие системы видотипичных, врожденных действий включаются воздействием внешних, экзогенных агентов, в нашем примере — действительным приближением врага, воспринимаемого экстероцепторами. В этом случае постепенное его приближение вызовет усиление раздражения соответствующих участков мозговых структур, что в эксперименте достигалось искусственно при помощи воздействий электрическим током.
Таким образом, поведение в своей основе внутренне так же организовано, так же зиждется на системах биологической саморегуляции, так же закодировано в генетическом фонде вида, как и процессы, определяющие другие функции организма. В этом и проявляется единство всех форм жизнедеятельности животных.

Внешние факторы инстинктивного поведения
Когда говорят об автономности внутренних факторов поведения, об их независимости от внешней среды, то необходимо помнить, что эта независимость является лишь относительной. Уже из приведенных опытов Хольста видно, что эндогенная активность не существует ни «сама по себе», ни «сама для себя»: значение этих спонтанных процессов в центральной нервной системе состоит в прениспозиции к возникновению жизненно важных ситуаций («в случае чего — все готово»). В результате животное способно по первому же сигналу незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде.
Эта готовность обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (например, изменением продолжительности светового дня, повышением или уменьшением температуры и т. д.). Однако инстинктивные движения, по этологической концепции, заблокированы специальной системой «врожденных пусковых механизмов». Последние представляют собой совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к «пусковой ситуации»). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие «блокировки». Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости.
Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию. Иными словами, в сенсорной сфере существует некая «фильтрующая» функция, выражающаяся в специфической преадаптационной «готовности» воспринимать такие раздражители.
Итак, благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает «выход наружу», т. е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида.
Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к «пусковой ситуации»: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом.
Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название «ключевых раздражителей», поскольку они подходят к своим врожденным пусковым механизмам, как ключ к замку. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видотипичными формами поведения, точнее, определенными инстинктивными движениями. В описанном поведении курицы это будут определенные общие признаки, свойственные всем ее наземным врагам.
Кроме собственно растормаживающих ключевых раздражителей (их называют также «пусковыми раздражителями») различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки («просто» форма, размер, подвижность, цвет, запах и т. д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т. д.), или же векторы. Носителями этих признаков могут быть как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, например, немецкий этолог Ф. Вальтер показал, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно), является «что-то вертикальное» вне зависимости от того, что конкретно это за объект.
Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н. Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция («попрошайничество») птенцов серебристых чаек (клевание клюва родительской особи) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи.
В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и тот в ответ отрыгивает пищу в рот птенцу. Об этом говорит сайт https://intellect.icu . В опытах предъявлялась серия все более упрощаемых моделей — макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т. е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб постепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 7). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но этот предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция может быть усилена, если этот макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто «красное» и «продолговатое».
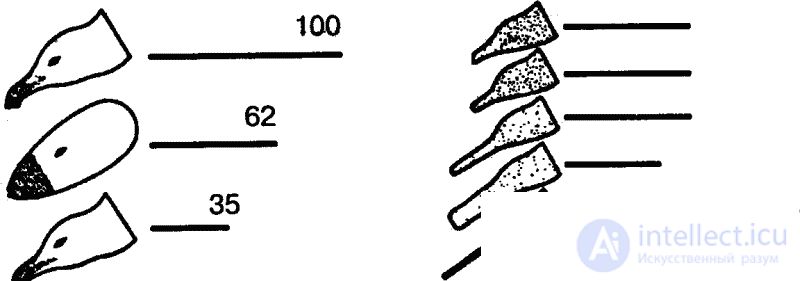
В опытах, проведенных Тинбергеном с десятидневными птенцами дроздов, выявилась другая категория ключевых раздражителей — взаиморасположение и относительная величина деталей объекта-носителя. В качестве макетов использовались плоские диски. Если показать птенцам такой круг, они будут тянуться к его верхней части (местонахождению головы птиц). Если же к большому кругу прибавить маленький, птенцы потянутся к нему. Если, наконец, прибавить к большому кругу два меньших круга, но разных размеров, то решающее значение приобретет относительная величина этих фигур. Размеры «головы» не должны приближаться к размерам «туловища»: при большом «туловище» птенцы потянутся к большему дополнительному кругу («голове») (рис. 8), при маленьком же—к меньшему.
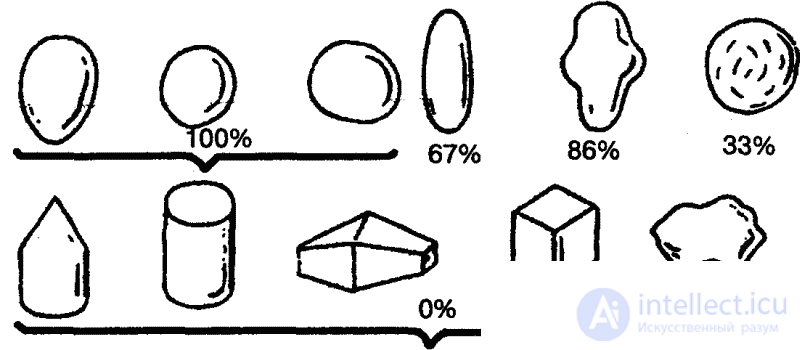
Интересные опыты проводил еще в середине 30-х годов Г. Л. Скребицкий совместно с Т. И. Бибиковой на подмосковном озере Киево, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, искусственными яйцами, а затем и различными посторонними предметами различной величины, формы и окраски. Оказалось, что чайки садились как на чужие, так и на деревянные, стеклянные, каменные, глиняные яйца различной величины и самой разнообразной окраски и начинали их «высиживать». То же самое наблюдалось, когда вместо яиц в гнездо подкладывались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, «чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку».[35]
Если, однако, чайкам подкладывались предметы иной формы, например кубики или камни с неровными краями, поведение птиц заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска.

Исследователи пришли к выводу, что положительная реакция чайки на яйцо определяется лишь несколькими его элементарными признаками: округлостью, отсутствием выступов, углублений или насечек (рис. 9). Именно эти признаки и выступали здесь как ключевые раздражители.
Важная особенность действия ключевых раздражителей заключается в том, что они подчиняются закону суммации: с увеличением их параметров пропорционально усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым «супероптимальным» реакциям, когда животное «преувеличенно», сильнее, чем в норме, реагирует на искусственный раздражитель, в котором «сгущены краски». Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности.
Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. Если, например, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя.
Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных.
Так, например, еще в начале нашего века один из авторов антропоморфических сочинений по психологии животных Т. Целл дал следующий ответ на вопрос о том, почему крупные хищники в нормальных условиях при встрече с человеком не нападают на него: например, лев не нападает не потому, что почитает человека, а потому, что не уверен в исходе схватки. «Есть ли у человека оружие, да какое оно, это оружие? — думает лев. — Да ну его, пойду-ка я лучше своей дорогой».
Конечно, лев, как и другие крупные хищники, как правило, уклоняется от нападения на человека не потому, что руководствуется такими глубокомысленными рассуждениями. Разгадка «почтительного» отношения диких зверей к человеку, очевидно, кроется в следующем. Будучи сытым, хищник не реагирует и на присутствие животных, которыми обычно питается. У голодного же хищника преследование животных-жертв и нападение на них обусловливаются сочетанием рассмотренных выше внутренних факторов (первопричина — изменение уравновешенности внутренней среды организма в результате недостатка питательных веществ) с соответствующими внешними стимулами — ключевыми раздражителями, носителями которых являются естественные объекты питания, т. е. животные-жертвы, но не человек.
Даже самый кровожадный хищник не волен нападать на кого угодно и когда угодно. Эти действия также ориентируются во времени и пространстве ключевыми раздражителями, как и все прочие поведенческие акты. Другими словами, дело не в том, «хочет» или «не хочет» животное поступить так или иначе. Если внутреннее состояние животного соответствует определенной внешней пусковой ситуации, то оно волей-неволей вынуждено вести себя так, как это диктует для данных условий генетически зафиксированный код видотипичного поведения.
Итак, в процессе эволюции возникают адаптации к более постоянным компонентам внешней среды, необходимым для удовлетворения потребностей, непрестанно возникающих в результате изменений внутренней среды организма. Нахождение (или избегание) важных для организма компонентов внешней среды осуществляется путем ориентации по типичным признакам этих компонентов — ключевым раздражителям.
Результаты этой ориентации реализуются нейросенсорными системами (врожденными пусковыми механизмами), которые действуют рефлекторно и включают эндогенные, генетически фиксированные компоненты инстинктивного поведения. Таким образом, действуя «вовне», врожденные пусковые механизмы обеспечивают избирательную направленность внешней активности организма лишь на определенные сигнальные стимулы; действуя же «вовнутрь», они осуществляют оценку и отбор поступающей через рецепторы информации и ее реализацию для активации или понижения порогов раздражимости соответствующих нервных структур, для снятия «блокировки», растормаживания эндогенных нервных процессов, мотивирующих инстинктивные движения и действия. Таким образом осуществляется на врожденной основе корреляция внутренних потребностей организма с биологически существенными изменениями в окружающей его среде.
Только в этой корреляции состоит все биологическое значение эндогенной мотивации поведения. Внутренние стимулы служат лишь для осуществления движения по отношению к среде, без которого организм — именно как саморегулирующаяся система — нежизнеспособен. И в этом, широком, смысле деятельность всей нервной системы в целом всегда рефлекторна.
Равным образом даже наиболее как будто «далеко упрятанные» от внешней среды факторы поведения сами зависят от процессов обмена веществ. Эти процессы уже непосредственно связаны с окружающей организм средой. А так как организм активно регулирует, создает необходимые внешние предпосылки для нормального протекания обменных процессов как раз посредством поведения, то круг смыкается. И в этом отношении проявляется относительность независимости эндогенных автоматизмов, спонтанной активности нервной системы, способности к «самопрограммированию». Ясно, что все эти эндогенные процессы лишь постольку независимы от внешней среды, поскольку они лишь опосредованно связаны с ней.
Структура инстинктивного поведения
Поисковая и завершающая фазы поведенческого акта
Выше говорилось о том, что ключевые раздражители действуют принудительно, что животное вынуждено в своем поведении всецело повиноваться пусковой ситуации. Но значит ли это, что у животных нет никакой возможности проявить собственную инициативу, произвести какой-то самостоятельный выбор? Отнюдь нет!
Инициативное, избирательное отношение животного к среде проявляется прежде всего в активном поиске необходимых пусковых ситуаций и в избрании наиболее эффективных возможностей для выполнения поведенческих актов. Необходимо подчеркнуть, что речь идет о поиске именно раздражений, исходящих от биологически значимых объектов, а не самих этих объектов. Мы теперь уже знаем, что это ключевые раздражители с направляющей или пусковой функцией.
Еще более полувека тому назад американский исследователь поведения животных У. Крэйг показал, что инстинктивные действия состоят из отдельных фаз. Прежде всего Крэйг выделил две фазы, которые в этологической литературе получили названия «поисковая» (или «подготовительная») и «завершающая». В ходе поисковой фазы животное отыскивает (отсюда и название фазы) те ключевые раздражители, точнее, их комбинации (т. е. пусковые ситуации), которые приведут его в итоге к завершающей фазе, в которой воплощается биологическое значение всего инстинктивного действия.
Все промежуточные раздражители не составляют для животного самоцель и ценны лишь постольку, поскольку они приводят к восприятию ключевых раздражителей завершающего поведения. Только на завершающей фазе происходит собственно потребление животным жизненно необходимых ему элементов среды. Но сам поиск адекватных раздражителей является для животных такой же первейшей жизненной необходимостью, как и потребление элементов среды.
Поисковая фаза всегда подразделяется на несколько этапов; однако, в завершающей же фазе, или вовсе не обнаруживаются такие подразделения, или же она состоит лишь из немногих строго последовательно выполняемых движений.
Крэйг построил свою концепцию на данных, полученных им в результате изучения пищевого поведения животных. Приведем пример этой сферы поведения. Хищник, отправляющийся на охоту, вначале еще не знает, где находится его возможная добыча, поэтому первые его движения имеют характер ненаправленного поиска. В результате он раньше или позже попадает в сферу действия стимула, исходящего от животного-жертвы. Обнаружен первый ключевой раздражитель, который включает следующий этап — направленную ориентировку по дополнительным раздражителям, уточнение местонахождения животного-жертвы. Затем следует подкрадывание (или преследование), набрасывание (прыжок) и овладение добычей, ее умерщвление, иногда еще и перетаскивание туши в другое место, расчленение на отдельные куски и, наконец, захватывание кусков мяса зубами и их проглатывание. В этой цепи последовательно выполняемых действий и движений только последние два звена (собственно акт поедания) относятся к завершающей фазе описанного пищедобывательного поведения хищника, все остальные этапы составляют в своей совокупности поисковое (или подготовительное) поведение. Правда, внутри каждого такого этапа имеются свои подготовительные и завершающие фазы, которыми каждый этап и заканчивается. При этом подчас имеется несколько степеней соподчинения (по типу «матрешки»), так что в целом складывается весьма сложная структура деятельности.
Аналогично обстоит дело и в других, казалось бы, значительно более простых сферах поведения, как, например, покой и сон. Животное вначале ищет место для отдыха или ночлега (деревья, укрытия, углубления в почве или просто определенные участки открытого пространства), затем устраивает (совершенствует) найденное место (копает, приминает растительность), иногда еще и чистится и только после этого укладывается (причем видотипичным образом!). Только укладывание составляет завершающую фазу, предшествующие же этапы — поисковую.
К этим примерам можно было бы прибавить еще множество других из любой сферы поведения. Однако уже в приведенных можно усмотреть следующие глубокие различия между обеими фазами, определяющие их сущность.
Поисковое поведение является пластичной фазой инстинктивного поведения. Она характеризуется ярко выраженной ориентировочно-исследовательской деятельностью животных и переплетением врожденных и приобретенных, основанных на индивидуальном опыте компонентов поведения. Именно к поисковому поведению относится все, что связано с пластичностью инстинктов, в частности с модификациями инстинктивного поведения.
Завершающее поведение, наоборот, представляет собой ригидную фазу. Выполняемые в ней движения отличаются строгой последовательностью, стереотипностью и предопределены соответствующими макро- и микроморфологическими структурами. Приобретенные компоненты играют здесь несущественную роль или даже отсутствуют. Поэтому изменчивость ограничена индивидуальной (генетически фиксированной) вариабельностью. Сюда относится все, что говорилось о постоянстве, ригидности инстинктивного поведения и о принудительности действия ключевых раздражителей. Здесь практически все врожденное, генетически фиксированное. Что же касается движений, выполняемых в завершающей фазе, то это и есть собственно инстинктивные движения, или «врожденные двигательные координации», названные так австрийским ученым К. Лоренцем, одним из основателей современной этологии.
Инстинктивные движения и таксисы
Общая характеристика инстинктивных движений уже давалась выше. Также говорилось о том, что они являются «хранителями» самого ценного, жизненно необходимого, что накоплено видом в результате естественного отбора, и что именно этим определяется их независимость от случайных условий окружающей среды. Лисица, производящая инстинктивные движения закапывания мяса на каменном полу, ведет себя «бессмысленно». Но ведь не на каменном субстрате совершался филогенез лисиц, и было бы фатально для дальнейшего существования вида, если бы из-за случайного, временного пребывания особи в таких совершенно нетипичных для местообитания лисиц условиях исчезла бы столь полезная для этих животных форма поведения. Так что лучше произвести закапывающие движения при любых обстоятельствах, и «жесткая» врожденная программа поведения заставляет животное это сделать.
Общая ориентация инстинктивных движений осуществляется таксисами, которые, по Лоренцу, всегда переплетаются с врожденными двигательными координациями и совместно с ними образуют единые инстинктивные реакции (или цепи из нескольких таких реакций).
Как и инстинктивные движения, таксисы — это врожденные, генетически фиксированные реакции на определенные агенты среды. Но если инстинктивные движения возникают в ответ на пусковые раздражители, то таксисы отвечают на направляющие ключевые раздражители, которые неспособны обусловливать начало (или конец) какой-либо инстинктивной реакции, а лишь меняют вектор ее протекания.
Таким образом, таксисы обеспечивают пространственную ориентацию двигательной активности животных в сторону благоприятных или жизненно необходимых условий среды (положительные таксисы) или же, наоборот, от биологически малоценных или опасных условий (отрицательные таксисы). У растений аналогичные реакции выражаются в изменениях направления роста (тропизмы).
По характеру ориентирующих внешних стимулов таксисы подразделяются на фото-, хемо-, термо-, гео-, рео-, анемо-, гидротаксисы (реакции на свет, химические раздражители, температурные градиенты, силу тяжести, течение жидкости, поток воздуха, влажность среды) и т. д. На разных уровнях эволюционного развития таксисы имеют разную степень сложности и выполняют различные функции, о чем еще пойдет речь при обзоре эволюции психики. Сейчас важно подчеркнуть, что таксисы являются постоянными компонентами даже сложных форм поведения, причем высшие формы таксисов выступают в тесном сочетании с индивидуальным опытом животного.
С направляющими ключевыми раздражителями мы уже встречались, когда описывалась ориентация птенцов по относительным оптическим стимулам. Само появление объекта (в эксперименте — диска, в естественных условиях — родительской особи) является пусковым раздражителем реакции «попрошайничества», взаиморасположение деталей этого объекта — направляющим ключевым раздражителем этой реакции, а пространственная ориентация птенцов по этому раздражителю — положительным фототаксисом.
Точно так же красный цвет сам по себе является для птенца серебристой чайки пусковым ключевым раздражителем, обусловливающим его пищевую реакцию (наряду с другим пусковым раздражителем — появлением птицы, точнее, ее головы с клювом). Местонахождение же красного пятна на клюве направляет реакцию птенца биологически выгодным образом на основе положительного фототаксиса и служит, таким образом, направляющим ключевым раздражителем.
Лоренц и Тинберген в 30-е годы совместно изучали взаимоотношения врожденных двигательных координации и таксисов на примере реакции вкатывания яиц в гнездо у серого гуся. Вид яйцеподобного предмета (нечто круглое, без выступов и т. д.), находящегося за пределами гнезда, служит у этой птицы таким же ключевым раздражителем для реакции вкатывания, как у сидящей на гнезде чайки в описанных опытах Скребицкого. Соответствующая врожденная двигательная координация представляет собой многократно повторяемое движение клюва к груди птицы, которое прекратится лишь тогда, когда предмет прикоснется к сидящей в гнезде птице.
Если положить перед краем гнезда перпендикулярно к клюву цилиндр (гуси положительно реагировали и на такой предмет), то подобными инстинктивными движениями и ограничится все поведение птицы. Если же положить яйцо или его макет, то появляются дополнительные движения головы из стороны в сторону, придающие движению предмета правильное направление к гнезду. Ведь в отличие от цилиндра яйцо будет откатываться то влево, то вправо. Вид этих отклонений и служит направляющим раздражителем для таксисных боковых движений головы. Итак, таксисы могут у высших животных ориентировать инстинктивные движения не только всего организма, но и отдельных частей тела и органов.
Из инстинктивных движений и таксисов складывается завершающая фаза каждого поведенческого акта. Вместе с тем они входят как составляющие части в поисковую фазу, которая, как уже говорилось, служит для поиска внешних пусковых ситуаций, позволяющих организму достигнуть завершающей фазы данного акта. Поисковая фаза отличается большой лабильностью и весьма сложной структурой. Инстинктивные движения завершают каждый промежуточный этап этой фазы, в результате конец каждого такого этапа также приобретает черты завершающего поведения. Таксисы дополняются в поисковой фазе ориентировочно-исследовательскими реакциями, непрерывно доставляющими организму информацию о состоянии, параметрах и изменениях компонентов окружающей среды, что позволяет ему произвести оценку последних в рамках общего поискового поведения.
Благоприобретаемые компоненты инстинктивного акта
Наряду с указанными в поисковой фазе любого инстинктивного акта всегда содержатся — в разной степени и в разных сочетаниях — и все элементы поведения, которые относятся к научению, не исключая и высших форм поведения, интеллектуального типа. Именно поэтому мы и считаем, что говорить об инстинктивных действиях (но не движениях!) — значит говорить о поведенческих актах вообще.
В самом деле, все, чему животные способны научиться, направлено лишь к одному — по возможности к скорейшему и наиболее экономному достижению завершающего поведения. Нет в поведении животных ничего, что бы не завершалось этой конечной фазой — инстинктивными движениями или родственными им реакциями, т. е. врожденными двигательными координациями. И в этом отчетливо видно единство инстинктивного поведения и научения.
Конечно, сказанное не относится в равной степени ко всем этапам поисковой фазы. Дело в том, что лабильность поведения неодинакова на разных этапах этой фазы и все больше уменьшается по мере приближения к завершающей фазе.
Переход от одного этапа поискового поведения к другому означает активный поиск и нахождение животными ключевых раздражителей, смену одного этапа поискового поведения другим в строго закономерной последовательности. Это сопровождается ступенчатым сужением сферы активности животного: его поведение все более детерминируется специфическими для данного поведенческого акта сочетаниями раздражителей, становится все более направленным в сторону конечных, завершающих инстинктивных движений, суживаются возможности индивидуального видоизменения поведения животного, пока завершающая пусковая ситуация не сведет эти возможности практически к нулю.
Так, например, ласточка, приступая к постройке гнезда, должна, прежде всего, найти место, где можно собирать гнездостроительный материал. Первоначально ненаправленный осмотр местности является первым этапом гнездостроительной деятельности. Скорость прохождения этого предельно лабильного этапа зависит, прежде всего, от индивидуально-изменчивых компонентов поведения, прежде всего от уже имеющегося индивидуального опыта. В зависимости от своих индивидуальных психических способностей каждая птица решает эту задачу по-своему, более или менее эффективно.
Следующий этап поисковой фазы — поиск и сбор гнездостроительного материала на найденном подходящем месте. Здесь возможности индивидуального изменения видотипичного поведения уже сужены, но тем не менее индивидуальная сноровка продолжает играть немаловажную роль, Еще меньше амплитуда возможных индивидуальных отклонений на основе прежнего опыта на третьем этапе поискового поведения — транспортировке гнездостроительного материала к месту постройки гнезда. Здесь возможны лишь некоторые, не очень существенные вариации в скорости и траектории полета. В остальном поведение всех ласточек уже весьма стереотипное.
И наконец, завершающая фаза — прикрепление частичек к субстрату — выполняется уже совершенно стереотипными инстинктивными движениями. Здесь только генетически обусловленная индивидуальная вариабельность видотипичного поведения дает уже известные нам различия в строении отдельных гнезд.
Таким образом, постепенно уменьшается «свобода действия» животного, уменьшается амплитуда изменчивости его поведения по мере приближения к завершающей фазе в соответствии со все более ограниченными и специфическими условиями среды, сочетаниями раздражителей, причастных к данному поведению.
Чем дальше от завершающей фазы и чем больше амплитуда изменчивости видотипичного поведения, тем больше возможностей включения в него элементов научения, индивидуального опыта и тем больше удельный вес этих элементов. Таким образом, индивидуальный опыт реализуется прежде всего на начальных этапах поискового повелении. И еще: чем выше психическое развитие, тем существеннее коррективы, вносимые в жесткое видотипичное поведение, но опять же главным образом на начальных этапах поисковой фазы. Все это относится, конечно, и к поисковому и завершающему поведению внутри каждого этапа поискового поведения.
Сложность и разнообразие структуры инстинктивного поведения
Двухфазная структура инстинктивных действий дана здесь лишь в виде весьма полной, упрощенной общей схемы. В реальной действительности чаще всего имеют место разные осложнения и видоизменения. Прежде всего необходимо иметь в виду, что поисковая фаза может происходить и под отрицательным знаком — в виде уклонения и избегания определенных агентов среды. Далее возможны сокращения поискового поведения, выпадения отдельных его этапов или даже инверсия. Иногда завершающие движения наступают так быстро, что поисковая фаза не успевает проявиться в полной мере. В других случаях поисковое поведение может свернуть со своего русла и привести к «чужому» завершающему поведению.
Поисковое поведение может принять форму завершающего поведения и существовать наряду с истинной завершающей фазой. В таком случае внешне одинаковые действия будут иметь двоякую качественно различную мотивацию.
Большой интерес представляют и различные случаи неполного протекания инстинктивного акта, когда действия животного не достигают завершающей фазы. У животных с наиболее высокоразвитой психикой промежуточные этапы поискового поведения, т. е. сам по себе поиск раздражителей, могут при этом — как исключение — стать самоцелью их поведения. Здесь мы встречаемся с инстинктивной основой наиболее сложных форм исследовательского поведения, входящих в фундамент интеллекта животных.
Уже этот перечень показывает все разнообразие протекания инстинктивных действий; добавим еще, что практически никогда в одно и то же время не совершается лишь единственный инстинктивный акт, а имеет место сложное взаимодействие между несколькими одновременно происходящими действиями.
Инстинктивное поведение и общение
Все животные периодически вступают во внутривидовые контакты друг с другом. Прежде всего это относится к сфере размножения, где часто наблюдается более или менее тесный контакт между половыми партнерами. Кроме того, представители одного и того же вида часто скапливаются в местах с благоприятными условиями существования (обилие пищи, оптимальные физические параметры среды и пр.). В этих и подобных случаях происходит биологическое взаимодействие между животными организмами, на основе которого в процессе эволюции зародились явления общения. Ни любое контактирование между самцом и самкой, ни тем более скопление животных в благоприятных для них местах (зачастую с образованием колонии) не является проявлением общения. Последнее, как и связанное с ним групповое поведение, предполагает как непременное условие не только физическое или биологическое, но прежде всего психическое взаимодействие (обмен информацией) между особями, выражающееся в согласовании, интегрировании их действий. Как еще будет показано, это относится к животным, стоящим выше кольчатых червей и низших моллюсков.
Дополняя сказанное, необходимо подчеркнуть, что об общении можно говорить лишь тогда, когда существуют особые формы поведения, специальной функцией которых является передача информации от одной особи к другой. Другими словами, общение, в научном значении этого термина, появляется лишь тогда, когда некоторые действия животного приобретают сигнальное значение. Немецкий этолог Г. Темброк, посвятивший много усилии изучению процессов общения и их эволюции, подчеркивает, что о явлениях общения и соответственно подлинных сообществах животных (стадах, стаях, семьях и т. д.) можно говорить лишь тогда, когда имеет место совместная жизнь, при которой несколько самостоятельных особей осуществляют вместе (во времени и пространстве) однородные формы поведения в более чем одной функциональной сфере. Условия такой совместной деятельности могут меняться, иногда она осуществляется при разделении функций между особями.
Добавим еще, что если общение отсутствует у низших беспозвоночных и только в зачаточных формах появляется у некоторых их высших представителей, то, наоборот, оно присуще всем высшим животным (включая и высших беспозвоночных), и можно сказать, что в той или иной степени поведение высших животных в целом осуществляется всегда в условиях общения (хотя бы периодического).
Как уже упоминалось, важнейшим элементом общения является обмен информацией — коммуникация. При этом информативное содержание коммуникативных действий (зоосемантика) может служить опознаванию (принадлежности особи к определенному виду, сообществу, полу и т. п.), сигнализировать о физиологическом состоянии животного (голоде, половом возбуждении и пр.) или же служить оповещению других особей об опасности, нахождении корма, места отдыха и т. д. По механизму действия (зоопрагматика) формы общения различаются каналами передачи информации (оптические, акустические, химические, тактильные и др.), но во всех случаях коммуникации животных представляют собой (в отличие от человека) закрытую систему, т. е. слагаются из ограниченного числа видотипичных сигналов, посылаемых «животным-экспедиентом» и адекватно воспринимаемых «животным-перцепиентом».
Общение между животными невозможно без генетической фиксации способности как к адекватному восприятию (что обеспечивается таксисами), так и к передаче «кодированной» информации (что обеспечивается врожденными пусковыми механизмами). Наследственно закрепленные, инстинктивные действия, с помощью которых выполняется передача информации, могут приобретать самостоятельное значение. Здесь действуют те же закономерности, что и при других формах инстинктивного поведения, но носителями ключевых раздражителей являются в данном случае сородичи или животные других видов.
Среди оптических форм общения важное место занимают выразительные позы и телодвижения, которые состоят в том, что животные весьма заметным образом показывают друг другу определенные части своего тела, зачастую несущие специфические сигнальные признаки (яркие узоры, придатки и т. п. структурные образования). Такая форма сигнализации получила название «демонстрационное поведение». В иных случаях сигнальную функцию выполняют особые движения (всего тела или отдельных его частей) без специального показа особых структурных образований, в других — максимальное увеличение объема или поверхности тела или хотя бы некоторых его участков (посредством его раздувания, расправления складок, взъерошивания перьев или волос и т. п.). Все эти движения всегда выполняются «подчеркнуто», нередко с «преувеличенной» интенсивностью (рис. 10). Но, как правило, у высших животных все движения имеют какое-то сигнальное значение, если они выполняются присутствии другой особи.
Наиболее полноценная, четкая передача информации достигается, однако, тогда, когда появляются специальные двигательные элементы, выделившиеся из «обычных», «утилитарных» форм поведения, утративших в ходе эволюции свою первичную «рабочую», «механическую» функцию и приобретшие чисто сигнальное значение.

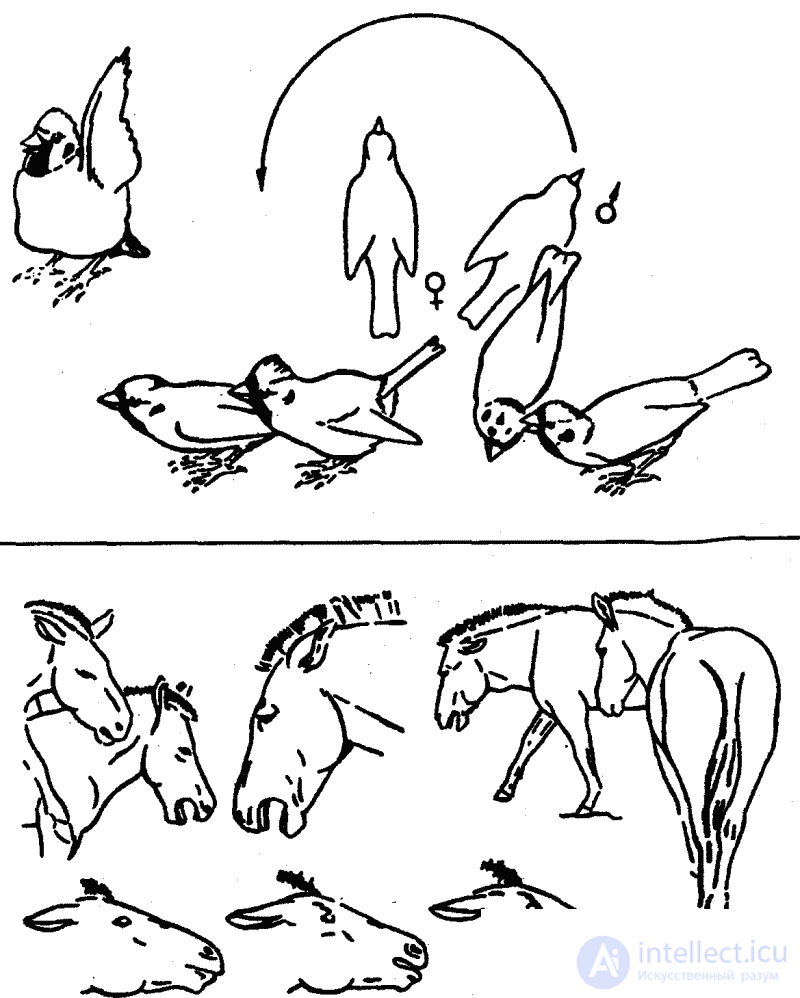
Первичные движения получили в этологии название «автохтонных» движений; вторичные, приобретшие новую, в данном случае сигнальную, функцию, — «аллохтонных». В таком случае сам рисунок стереотипно, у всех представителей данного вида одинаково выполняемого движения является условным выражением определенной биологической ситуации (биологически значимого изменения в среде или внутреннего состояния животного). Такие видотипичные стереотипные движения с четкой информативной функцией получили в этологии название «ритуализованных движений». Ритуализация характеризуется условностью выполняемых действий, которые служат лишь для передачи определенного, причем совершенно конкретного информативного содержания. Эти действия генетически жестко фиксированы, выполняются с максимальной стереотипностью, т. е., по существу, одинаково всеми особями данного вида, и поэтому относятся к типичным инстинктивным движениям. По этой причине все животные данного вида и в состоянии «правильно» понимать значение подобных оптических (или акустических) сигналов общения. Это типичный пример максимальной ригидности, консерватизма в поведении животных.
Ритуализованные движения выполняются в строгой последовательности в форме более или менее сложных видотипичных «церемониалов» или «ритуалов», причем, как правило, в виде «диалога» двух животных. Чаще всего они встречаются в сферах размножения (брачные игры) и борьбы («мнимая борьба») и отображают внутреннее состояние особи и ее физические и психические качества. Наряду с врожденными существуют и благоприобретаемые формы общения, о которых пойдет речь позже.
Все формы общения играют существенную роль в жизни высших беспозвоночных и позвоночных, обеспечивая согласованность действий особей. Без коммуникации с помощью различных химических, оптических, акустических, тактильных и других сигналов у этих животных невозможно даже сближение и контактирование самцов и самок, а, следовательно, и продолжение рода. Особое значение приобретает общение особей одного, реже разных видов в условиях их совместной, групповой жизни как основа группового поведения.
Как уже говорилось, групповое поведение появляется (по меньшей мере в развитых формах) вместе с общением только у высших беспозвоночных и выражается в согласованных совместных действиях животных, живущих в сообществах. В отличие от скоплений животных, возникающих в результате идентичной положительной реакции на благоприятные внешние условия, сообщества характеризуются относительно постоянным составом его членов и определенной внутренней структурой, регулирующей отношения между ними. Эта структура нередко принимает форму иерархических систем соподчинения всех членов сообщества друг другу, чем обеспечивается сплоченность последнего и его эффективное функционирование как единого целого. Не меньшее значение имеет в этом отношении выделение вожаков, или «лидеров», регулирование отношений между молодыми и старыми животными, совершенствование ухода за потомством и т. д.
Психический компонент инстинктивного поведения
Психический компонент инстинктивного поведения, точнее, психическое отражение при инстинктивных действиях животных необходимо изучать на завершающей фазе поведенческого акта и в какой-то степени на непосредственно примыкающем к ней последнем этапе поисковой фазы, поскольку здесь элементы научения играют наименьшую роль. При этом надо сказать, что в свете современных данных и концепций этологии этот вопрос еще специально не изучался и поэтому приходится здесь ограничиваться лишь некоторыми общими соображениями.
Мы видели, что по мере приближения к завершающей фазе движения животного становятся все более ограниченными, однообразными, стереотипными, и есть все основания полагать, что в такой же мере отражение окружающего мира становится все более бедным, ограниченным. Если на первых этапах поискового поведения животное в поиске ключевых раздражителей руководствуется многими разнообразными ориентирами в окружающей среде, то по мере сужения действия внешних агентов, направляющих поведение животного, ориентация все больше осуществляется только по направляющим ключевым раздражителям, а в завершающей фазе производится уже исключительно по ним.
Вместе с тем мы теперь уже знаем, что как направляющие, так тем более и пусковые ключевые раздражители представляют собой элементарные физические и химические признаки, воспринимаемые животным «в отрыве» от объекта, которому они присущи. Следовательно, животное получает на завершающей фазе лишь весьма неполную, «однобокую» информацию о некоторых — причем несущественных — внешних признаках объектов инстинктивных действий. Правда, эти признаки являются постоянными и в этом отношении характерными для этих объектов.
Однако, что для нас сейчас важно, животное, реагируя на эти признаки, в сущности, не получает сведений о самом объекте (неодушевленном предмете или другом живом существе): признаки — ключевые раздражители — являются только ориентирами, направляющими действия животного на этот объект, носитель этих ориентиров.
Это с совершенной очевидностью вытекает из приведенных выше экспериментов, в которых подлинные объекты инстинктивных действий заменялись весьма абстрактными макетами. Вспомним, что в этих условиях удалось получить даже сверхоптимальные «преувеличенные» инстинктивные реакции. Самец колюшки в завершающей фазе борьбы за гнездовой участок не видит разницы между живой рыбой-соперником и куском жести или пластмассы. Он видит только «красное», и этого ему в данной ситуации вполне достаточно, чтобы среагировать на это «красное», которое в данном случае, кстати, выступает одновременно как направляющий и пусковой ключевой раздражитель.
Следовательно, здесь налицо первично генерализованная реакция на общий элементарный признак (точнее, на совокупность нескольких таких признаков, поскольку в природных условиях действуют не одно только «красное», но и некоторые другие ключевые раздражители). Этот признак воспринимается недифференцированно, как простое ощущение (точнее, воспринимаются суммы ощущений). Об этом свидетельствует отсутствие реакций на форму объекта — носителя признака, да и вообще на объект (предмет или рыбу) как таковой. Ясно, что здесь обнаруживается очень бедное, крайне поверхностное и ограниченное отражение окружающего мира, низшая форма психического отражения. Неинтересно, что такое отражение, свойственное низшим представителям животного мира, в полной мере сохранилось и у высших животных — позвоночных и продолжает у них играть главную роль в поддержании жизнедеятельности особи и продолжении рода.
О том, как это понимать, уже говорилось выше. Эволюция животной жизни могла идти только по пути отбора как можно более простых, но и наиболее постоянных ориентиров (или, что не меняет существа дела, создания таких специфических ориентиров, когда речь идет о животных одного вида, являющихся носителями ключевых признаков), руководствуясь которыми организмы могли бы успешно, не отвлекаясь другими, случайными, изменчивыми признаками, осуществлять наиважнейшие жизненные функции.
Правда, как явствует, например, из описанных опытов с птенцами дроздов, у высших животных ключевые стимулы представлены и более сложными по своей структуре типами. В указанном примере имеет место реакция на конфигурацию и на пространственные соотношения деталей объектов инстинктивных движений, что уже соответствует элементарным восприятиям. Это, очевидно, одно из важных проявлений прогрессивного развития инстинктивного поведения в ходе эволюции. Однако, как показывают те же эксперименты, и здесь нет подлинного предметного восприятия, ибо иначе птенцы не могли бы спутать свою мать с картонным диском.
Примитивность психического отражения на завершающей фазе инстинктивных действий является следствием бедности самой моторики в этой фазе. Как мы знаем, двигательная активность, направленная на окружающую среду, является источником познания этой среды. Однако столь стереотипные движения со столь ограниченной, специальной функцией, какими являются инстинктивные движения, врожденные двигательные координации, не могут служить сколько-нибудь пригодной основой для познания окружающего мира.
Врожденные двигательные координации (во всяком случае в их типичной форме) и не выполняют эту функцию, а ключевые раздражители, воспринимаемые на завершающей фазе (да и на отдельных этапах поискового поведения), не познаются, а узнаются врожденным образом благодаря наличию врожденных пусковых механизмов, чтобы затем использоваться как ориентиры или — и того меньше — как «кнопки» пуска инстинктивных реакций. Здесь больше ничего нет, кроме, вероятно, положительной или отрицательной эмоциональной окраски ощущений от воспринимаемых стимулов и собственных движений.
Познавательные же функции и вообще все богатство психического отражения приурочены к начальным этапам поискового поведения, где в полной мере действуют процессы научения.
Если я не полностью рассказал про инстинктивное поведение? Напиши в комментариях Надеюсь, что теперь ты понял что такое инстинктивное поведение и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Сравнительная психология и Зоопсихология

Комментарии