Лекция
Привет, мой друг, тебе интересно узнать все про развитие психической деятельности в раннем постнатальном периоде, тогда с вдохновением прочти до конца. Для того чтобы лучше понимать что такое развитие психической деятельности в раннем постнатальном периоде , настоятельно рекомендую прочитать все из категории Сравнительная психология и Зоопсихология.
Особенности постнатального развития поведения животных
Пре- и постнатальное развитие психической деятельности
Психическая деятельность эмбриона — это поведение и психика в процессе их становления на начальной стадии существования особи. Эмбрион является всего лишь формирующимся организмом, еще не способным к полноценному осуществлению функций, необходимых для установления жизненно важных взаимоотношений со средой обитания вида. В такой же степени и психика существует на пренатальной стадии развития лишь в зачаточной форме.
Рождение является поворотным пунктом, где весь процесс развития поведения получает новое направление. Естественно, что на уровне постнатального развития появляются совершенно новые факторы и закономерности, обусловленные взаимодействием организма с подлинной внешней средой. В этих новых условиях, однако, продолжается созревание врожденных элементов поведения и их слияние теперь уже с постнатальным, индивидуальным опытом. Поэтому, несмотря на коренные различия в условиях формирования пре- и постнатального поведения, между этими этапами онтогенеза поведения не только нет разрыва, но существует прямая преемственность. Именно в этом проявляется преадаптивное значение эмбрионального поведения, о чем также говорилось выше.
Если взять, к примеру, птиц, то, как было показано, к моменту вылупления уже существует достаточно развитая система общения между птенцом и высиживающей особью. Кроме того, существует акустическое общение и между невылупившимися птенцами внутри кладки, что влияет непосредственно на процесс вылупления. Так, у виргинской куропатки и японского перепела звуки, подаваемые одними птенцами, стимулируют активность других. В результате вылупление происходит дружно в течение приблизительно одного часа. Экспериментально этот эффект был доказан путем подкладывания яиц, находящихся на более ранней стадии инкубации, к яйцам, в которых находились птенцы перед самым вылуплением, что приводило к ускорению вылупления «отставших» птенцов на несколько часов. Акустическая коммуникация приводит, следовательно, у еще не вылупившихся птенцов к синхронизации вылупления. Преемственность же между эмбриональным и постэмбриональным поведением выражается в данном случае в том, что птенцы одного выводка, равно как птенцы и родители, отчасти уже «знают» друг друга как конкретных особей по индивидуальным признакам еще до появления первых на свет. Другие примеры, иллюстрирующие преадаптивное значение пренатального поведения, приводились в предыдущей главе.
Ранний постнатальный период имеет особенно большое значение для жизни особи, так как на этом этапе развития формируются важнейшие взаимоотношения организма с окружающей средой, устанавливаются связи с жизненно важными компонентами этой среды и закладываются основы поведения взрослого животного. Орбели говорил по этому поводу, что ранний постнатальный период является особо чувствительной фазой, наиболее жадно воспринимающей влияния, имеющие большое значение для дальнейшего формирования особи.
Разнообразие постэмбрионального развития поведения животных
Говоря о постнатальном развитии поведения животных, необходимо прежде всего иметь в виду, что оно протекает по-разному у разных животных и отличается специфическими закономерностями. Особенно это относится к тем животным, у которых новорожденные и ювенильные (т. е. молодые) формы резко отличаются строением и образом жизни от взрослых форм. У большинства беспозвоночных, а также ряда низших позвоночных личинки совершенно не похожи на взрослые особи, и зачастую им свойственны совершенно иные способы передвижения, питания и т. д.
Так, например, у морского моллюска из рода Strombus, как и у других морских брюхоногих, молодь ведет первоначально планктонный образ жизни, однако сразу же после опускания на дно и метаморфоза (т. е. превращения из одной стадии развития в другую) появляются свойственные взрослым животным формы придонной локомоции и питания. Вместе с тем защитная реакция — уход от хищника (хищных моллюсков рода Conus) — появляется только спустя три недели после метаморфоза, причем также сразу в «готовом виде». Предшествующий опыт (в эксперименте — контакт с водой, в которой содержался хищник) не ускоряет созревания этой реакции и не влияет на выраженность ее проявлений. В указанный же срок молодой Strombus тут же адекватно реагируют на приток воды, в которой находился его враг. Таким образом, мы имеем здесь дело со случаем весьма консервативного поведения, в той же мере генетически обусловленным, как и морфологические признаки. Постепенное усложнение поведения идет здесь строго «в ногу» с усложнением нервных структур и общего строения моллюска.
Аналогичные примеры известны также по членистоногим и другим беспозвоночным. Во всех этих случаях постэмбриональное развитие поведения совершается особенно сложным путем, ибо происходит резкая смена форм поведения и как бы в недрах личиночного поведения созревает качественно отличное взрослое поведение. Так, в приведенном примере личинка Strombus плавает на протяжении первой постэмбриональной стадии — в составе планктона — в поверхностных слоях моря, а затем переходит к придонному образу жизни и передвигается по субстрату характерными «прыжками».
Однако и в тех случаях, когда между ювенильными и взрослыми животными не обнаруживаются столь глубокие различия, филогенетические и экологические детерминанты вносят большое разнообразие в протекание постнатального онтогенеза поведения. Особенно большое значение имеет при этом степень зрелорождения потомства.
Постэмбриональное развитие поведения у зрело- и незрелорождающихся позвоночных
Детеныши позвоночных рождаются на разных стадиях зрелости. В предыдущей главе мы уже познакомились с примером крайнего незрелорождения — кенгуренком, появляющимся на свет еще в «полузародышевом» состоянии. Незрелорожденными, хотя и на более поздней стадии развития, являются птенцы многих птиц (воробьиных, хищных и др.), детеныши большинства млекопитающих (грызунов, хищных и др.). С другой стороны, птенцы кур, уток, гусей и ряда других птиц, как и детеныши копытных, представляют собой пример зрелорождения. В этом случае новорожденный почти сразу же, во всяком случае очень скоро, становится способным к самостоятельному выполнению основных жизненных функций (опорно-локомоторных, пищедобывательных и др.). Незрелорождающиеся же детеныши на протяжении определенного времени выполнять эти функции еще не могут и всецело зависят от родительского ухода. Промптов говорил в этом случае о «раннем биостарте». Под биостартом он понимал начало влияния биологических факторов на развитие птенца. Крайние проявления зрелорождения встречаются, правда, сравнительно редко. Чаще всего наблюдаются разные переходные степени зрелорождения. Например, у турухтана (отряд кулики) вылупившиеся птенцы проявляют много признаков зрелорождения как в рецепторной, так и в моторной сфере. В первые часы жизни они уже способны самостоятельно двигаться, затаиваться, вскоре они становятся способными и самостоятельно питаться. Вместе с тем они до 10-дневного возраста нуждаются в обогреве, как незрелорождающиеся птенцы, и координация движений вполне устанавливается лишь к 4-м суткам жизни. К этому сроку относится и полное формирование пищедобывательного поведения.
Орбели дал четкую биологическую характеристику развития поведения у зрело- и незрелорождающихся животных. Он указал на то, что, поскольку зрелорождающиеся детеныши начинают подвергаться воздействию среды в уже почти вполне сформированном состоянии, они сравнительно мало подвержены вредным влияниям среды. Но при этом чрезвычайно ограничены возможности дальнейшего прогрессивного развития поведения: имеют место лишь отдельные дополнительные надстройки условнорефлекторной деятельности.
Совершенно иное положение у животных, у которых развитие даже наследственно фиксированных, врожденных форм поведения выносится далеко за пределы внутрияйцевого или внутриматочного периода, т. е. у незрелорождающихся. По Орбели, эти детеныши, хотя им приходится труднее и они не могут жить без родительской помощи, находятся все же в более выгодном положении: развитие нервной системы у них еще не вполне закончено, и они могут подставить свои еще развивающиеся врожденные формы поведения под влияние агентов окружающей среды. В результате эти формы поведения в значительной степени модифицируются на основе переплетения врожденных и приобретаемых компонентов в соответствии с конкретными условиями среды. Как пишет Орбели, эти животные «родятся с настолько еще малосформированной нервной системой, что все дальнейшее постнатальное развитие представляет собою сплошную переработку наследственных форм и вновь возникающих условных форм поведения».[46]
С этой точки зрения более прогрессивными следует, безусловно, считать незрелорождающиеся виды животных, так как им начиная уже с «биостарта» обеспечено более пластичное, лабильное поведение, т. е. большая приспособляемость к постоянно меняющимся условиям среды. Правда, к кенгуру это не относится, так как здесь имеется, по существу, лишь кратковременный перерыв эмбриогенеза, как бы переход из одной утробы в другую, и ранняя смена способа питания — ведь в сумке матери крайне недоразвитый детеныш также весьма изолирован от внешнего мира и защищен от его вредных воздействий. (Важное отличие составляет, правда, возможное нахождение в той же сумке старшего детеныша.)
В этой связи интересно отметить, что психически наиболее развитые животные (а также человек, как подчеркивает и Орбели) в большинстве относятся к незрелорождающимся. У них новорожденный очень рано попадает в «настоящую» внешнюю среду, и генетически фиксированные формы поведения претерпевают здесь глубокую и длительную переработку. Правда, это не единственное обстоятельство, определяющее высокий уровень психического развития, ибо, например, гуси или лошади, т. е. типично зрелорождающиеся животные, также обнаруживают этот уровень.
По-видимому, решающее значение здесь имеют генетически обусловленные возможности накопления индивидуального опыта и его переноса в новые ситуации, активные воздействия на окружающую среду на основе развития эффекторно-рецепторных систем, двигательной активности, а в этом отношении различия между зрело- и незрелорождающимися животными сводятся к различиям в сроках формирования и развертывания этих систем и активности.
Что же касается этих сроков, т. е. общей скорости постнатального развития поведения, то они не отражаются на его результатах. Продолжительное постнатальное созревание, достройка и перестройка врожденного поведения, не влечет за собой замедления, задержки или тем более застоя общего развития поведения и наоборот. Японский орнитолог Н. Курода сопоставлял у разных видов воробьиных продолжительность гнездового периода и соответствующих форм гнездового поведения со сроками наступления способности к самостоятельному добыванию пищи у молодых птиц. Оказалось, что эта способность появляется приблизительно в одинаковые сроки независимо от длительности гнездового периода. Таким образом, при удлинении или укорочении гнездового периода птенец ничего не теряет, а в первом случае он быстро наверстывает «упущенное время» и оказывается не менее подготовленным к самостоятельной жизни, чем птицы, которые раньше покидают гнездо. Если учесть, что между зрело- и незрелорождением существуют переходные формы, то становится понятным, что и отмеченные различия имеют лишь относительное значение, тем более что темп онтогенеза на разных этапах меняется.
Значение заботы о потомстве
Большое значение, особенно у незрелорождающихся животных, приобретает родительская забота о потомстве, т. е. действия животных, обеспечивающие или улучшающие условия выживания и развития потомства. В процессе эволюции у многих групп животных возникли приспособления к защите и питанию развивающегося потомства со стороны родительской особи. Сюда относится и прохождение эмбриональных стадий развития в теле матери. Однако понятие «забота о потомстве» применяется лишь к постэмбриональному периоду. В ряде случаев забота о потомстве ограничивается созданием убежища и заготовкой пищи для будущего потомства, но материнская особь при этом не встречается с ним (превентивная забота о потомстве). Так, некоторые осы откладывают яйца на парализованных ими насекомых, которых прячут в специально вырытых норках, но затем уже не заботятся о вылупившихся личинках.
Более высокой формой заботы о потомстве является уход за потомством, проявляющийся в двух основных формах: пассивной и активной. В первом случае взрослые особи носят с собой яйца или молодых животных в специальных кожных углублениях, складках, сумках. Молодые животные при этом иногда питаются выделениями материнской особи. Эта форма ухода за потомством встречается у отдельных видов иглокожих, ракообразных, моллюсков, пауков, рыб (морской конек и игла, некоторые тропические окунеобразные — цихлиды), земноводных (жаба-повитуха, американская пипа, лягушка gastrotueca marsupiata), низших млекопитающих (ехидна, сумчатые). При активном же уходе за потомством взрослые особи выполняют специфические действия, направленные на обеспечение всех или многих сфер его жизнедеятельности — личинок насекомых, молоди рыб, птенцов, детенышей млекопитающих. Кроме устройства убежищ, кормления, обогрева, защиты, очищения поверхности тела и т. п. родители у многих высших животных (птиц и млекопитающих) также обучают свое потомство (например, находить пищу, распознавать врагов и т. д.).
Именно активный уход за потомством, высокоразвитая забота о нем делают возможным незрелорождение, а тем самым и все обусловленные им особенности психического развития. При этом эволюция заботы о потомстве знаменовалась, с одной стороны, интенсификацией и дифференциацией действий родителей по отношению к потомству, с другой стороны, усилением его зависимости от взрослых животных. Одновременно резко понизилась плодовитость. Однако возрастающая забота о потомстве влечет за собой и растущее противоречие между потребностями родительской особи и ее потомства. Это противоречие регулируется естественным отбором в сторону наибольшего прогресса вида. В. А. Вагнер охарактеризовал это формулой: минимум жертв матери — максимум требований потомства.
Таким образом, прогрессивные эволюционные приобретения, обеспечившие более гибкое приспособление растущего организма к условиям его жизни в постнатальном онтогенезе, имеют весьма сложную природу и включают в себя разные формы заботы о потомстве в зависимости от степени зрелорождаемости. Всем комплексом этих факторов определяется в каждом случае конкретный ход постнатального развития поведения.
Инстинктивное поведение в раннем постнатальном периоде
Инстинктивные движения
Стабильные, жесткие инстинктивные движения появляются в раннем постнатальном онтогенезе в настолько «готовом» виде, что долгое время считалось, что они совершенно не развиваются и не нуждаются в индивидуальном упражнении. На самом деле, как мы уже знаем, вполне врожденной является лишь «программа» их формирования в ходе онтогенеза. Сам же этот процесс обусловлен развитием экзосоматических эффекторных органов (и соответственных центральнонервных структур), функцией которых инстинктивные движения и являются. Только в таком смысле можно говорить о не зависящем от влияний среды появлении или созревании инстинктивных движений в онтогенезе.
Поскольку эти вопросы уже обсуждались раньше, мы ограничимся здесь лишь несколькими дополнительными замечаниями. Формирование инстинктивных движений в онтогенезе выявляется экспериментально путем изолированного выращивания детеныша с момента рождения (по методу «Каспар Гаузер»). Опыты, проведенные на птицах и млекопитающих (преимущественно грызунах), показали, что хотя поведенческие акты таких животных обнаруживают существенные отклонения от нормы, моторные элементы выполняемых при этом движений остаются без изменений, несмотря на то что детеныши были лишены возможности упражнения. (Курода, например, определил у птенцов серого скворца около 50 подобных элементов поведения, появляющихся спонтанно в разные сроки раннего постэмбрионального развития.) Это относится к особенно важным для жизни движениям, связанным с локомоцией, едой, спариванием, комфортными действиями и т. д. Но речь идет только о двигательных элементах, из которых слагаются эти движения. Длительность, частота, направленность, координированность, «отработанность» и совершенство этих движений, не говоря об их согласованности с другими поведенческими актами, а тем более с действиями других животных, оказывались в большей или меньшей степени нарушенными.
Большую известность приобрели проведенные американским ученым Э. Гессом опыты с цыплятами, которые вылуплялись в темноте, а затем были снабжены призматическими очками, смещавшими поле зрения на 7 градусов вправо. На 4-й день сопоставление с контрольными птенцами (снабженными очками, не искажавшими зрение) показало, что цыплята обеих групп клевали по целевому объекту (шляпке гвоздя) более точно, чем в первый день, но при этом не изменилось общее направление ударов: контрольные животные клевали по-прежнему на 7 градусов вправо от цели. Это доказывает, что данная двигательная координация, клевание и общее направление этих движений в поле зрения являются врожденными и не зависят от индивидуального опыта. Последний, однако, необходим для «отработки» точности ударов по цели, для обеспечения эффективности этой реакции.
Аналогично обстоит дело и у млекопитающих. Даже у обезьян было установлено, что моторные элементы двигательных реакций развиваются у новорожденных и без афферентной обратной связи (в эксперименте над макаками и павианами — после деафферентации передних конечностей у ослепленных детенышей). Вместе с тем, как показали, например, американский исследователь Г. Харлоу и его сотрудники, у макак-резусов и шимпанзе, выращенных в изоляции, появляются некоторые двигательные элементы нормального полового поведения, однако эти элементы не интегрируются в единый поведенческий акт, а в результате сексуальное поведение этих животных становится ненормальным. Аналогичные данные были получены и у морских свинок.
Таким образом, постнатальное развитие инстинктивных движений обнаруживает более или менее четкую генетическую обусловленность, не зависящую от раннего опыта. Но это относится только к элементарным врожденным двигательным координациям, но не к целым инстинктивным двигательным актам.
Врожденное узнавание
Появившись на свет, животное должно быстро и верно ориентировать свои движения по отношению к разным агентам среды. Такая ориентация по отношению к жизненно важным компонентам среды с минимальной тратой энергии и времени — поистине вопрос жизни и смерти, особенно для зрелорождающихся животных. Осуществляется она в первую очередь путем так называемого врожденного узнавания.
Врожденное узнавание (по Фабри) проявляется во врожденном, не зависящем от индивидуального опыта видоспецифическом избирательном отношении животных к определенным компонентам окружающей среды, признакам объектов, ситуациям, в способности животных биологически адекватно реагировать на некоторые признаки еще незнакомых им объектов или ситуаций. Таким образом, мы имеем здесь дело с врожденной формой ориентации, проявляющейся в полезных для особи (и вида) реакциях на признаки существенных компонентов среды без предварительного научения, с проявлениями «видовой памяти».
Подобные видотипичные положительные или отрицательные реакции животных, еще не успевших накопить индивидуальный опыт, направлены на такие компоненты среды, как, например, пища, материнская особь, особи своего вида; враги (или ситуации тревоги, даже если враг непосредственно не воспринимается) и т. п. Словом, речь идет о некоем преадаптивном узнавании объектов видотипичного инстинктивного поведения, позволяющего животному опознать биологически значимые объекты по некоторым их элементарным специфическим признакам без предварительного научения.
Основу такого опознавания составляют таксисы, но далеко не всегда здесь действуют простые безусловные рефлексы. Как показал Лоренц, во многих случаях подобная специфическая направленность поведения животного основана на соответствующих врожденных пусковых механизмах; сами же признаки, ориентирующие поведение животного, представляют собой ключевые раздражители.
Таким образом, оказавшись в биологически значимой ситуации, животное как бы узнает то, что никогда еще не воспринималось, с чем еще не приходилось познакомиться. Имеет место предварительная готовность к встрече с самыми важными для выживания элементами среды, т. е. как бы предваряется возможный будущий опыт особи.
Примеров врожденного узнавания можно привести множество. Напомним вновь о поведении новорожденного кенгуренка — его безошибочном нахождении сумки по ряду признаков, с которыми ему до этого не приходилось сталкиваться и в распознавании которых он не мог упражняться. Птенцы дроздов уже в первый день после вылупления, еще будучи слепыми, узнают о прилете родителей (с кормом) по легкому сотрясению гнезда и отвечают на это вытягиванием шеи и раскрытием клюва. Птенцы зреловылупляющихся птиц реагируют на вид подвижного объекта над головой припаданием к земле, что является эффективной защитой от воздушных врагов — хищных птиц. Крольчата в первый же день после рождения ориентируются в пространстве по тактильным раздражениям и таким образом находят источник пищи — соски матери. На каждое легкое прикосновение они реагируют движением в сторону раздражителя: при раздражении переднего конца головы перемещаются вперед, при прикосновении к спине или хвосту поворачиваются назад. Поворотами в сторону прикосновения реагируют также щенки и другие новорожденные млекопитающие.
Во всех случаях врожденного узнавания имеет место реакция на отдельные простые признаки биологически значимых объектов или же на сочетания или соотношения таких признаков, что закономерно и для реакции на ключевые стимулы. Имеет место и специфическая врожденная готовность к ощущению этих стимулов, характерная для врожденных пусковых механизмов. Всем этим и определяется избирательность врожденного узнавания, ее направленность лишь на определенные компоненты среды.
Эта отличительная особенность врожденного узнавания, как и вообще все связанные с ним закономерности, по Фабри, непосредственно обусловлена развитием врожденной двигательной активности. Как всегда, психическое развитие совершается и здесь в ходе и в результате развития моторики: врожденное узнавание возможно лишь как результат определенного уровня и специфической направленности развития врожденных компонентов моторики, о которых только что говорилось, т. е. тех специфических элементов движений, из которых слагается целостная реакция животного. При этом важно подчеркнуть, что речь идет именно о врожденных компонентах двигательной активности, появляющихся (или созревающих) в ходе раннего постнатального онтогенеза без научения.
Поясним сказанное примером. В лаборатории А. Д. Слонима осуществлялось с первого дня жизни перекрестное воспитание детенышей двух видов грызунов: большой песчанки (роющая форма) и белой крысы (нероющая форма). Независимо от этих условий молодые песчанки с 17-го дня жизни начинали проявлять типичную для их вида роющую деятельность, в то время как у крысят, которые воспитывались песчанками, роющие движения намечались в весьма слабом виде и то лишь начиная с 52-го дня жизни.
О чем говорят результаты этих опытов? Совершенно очевидно, что лишь после того как сформировались соответствующие двигательные способности конечностей, т. е. после того, как независимо от внешних условий воспитания появились врожденные, инстинктивные роющие движения, наступило врожденное инстинктивное узнавание рыхлого материала (песка, который ежедневно предъявлялся наряду с деревянными палочками в течение 10–15 минут) как возможного объекта рытья. Следовательно, избирательное узнавание и выбор этого материала по признаку рыхлости (ключевой раздражитель!) обусловлены необходимостью выполнения определенных движений, осуществления одной из жизненно важных для данного вида функций конечностей.
Врожденное узнавание и ранний опыт
В начале постнатального развития животного недопустима «роскошь» продолжительного научения. Тем не менее уже здесь проявляется общее правило, что нет чисто врожденных форм поведения, лишенных каких бы то ни было элементов научения. Это всецело относится и к врожденному узнаванию, которое всегда обогащается, корректируется или перестраивается в результате приобретения раннего опыта, проявляющегося в разных формах постнатального научения.
Так, нередко встречаются явления привыкания к часто повторяющимся биологически малозначащим стимулам. Птенцы куриных птиц, первоначально одинаково реагирующие на каждый подвижный объект, появляющийся над ними, вскоре научаются распознавать безопасные объекты (рис. 20). Это происходит путем привыкания: после неоднократного притаивания в ответ на появление в поле зрения безобидного объекта (листа, безопасной птицы и т. п.) эта реакция ослабевает и наконец совсем исчезает. Однако привыкает птенец лишь к объектам, которые он видит часто. В отношении же редких объектов необычного вида (а к таким и относятся хищные птицы с их характерным силуэтом) реакция, основанная на врожденном узнавании подвижного объекта в воздухе над головой, а именно затаивание, сохраняется в прежнем виде. Таким образом, мы имеем здесь дело с уточнением, дифференцировкой врожденного узнавания путем приобретения раннего опыта. Результатом этого процесса является в данном случае дифференцированное распознавание безопасных объектов.
Сходным образом, как уже указывалось, обстоит дело и у рыб, мальки которых на основе врожденного узнавания первоначально уплывают от любого крупного объекта или тени, надвигающихся на них. Впоследствии же они перестают реагировать на безопасные объекты, но научаются распознавать отличительные признаки хищника путем наблюдения за поимкой и поеданием членов стаи.

В результате раннего научения может меняться и сигнальное значение раздражителей. Молодь осетровых рыб реагирует первоначально на свет отрицательно, но начиная с 5-го дня (у шипа) или 9-го дня (у осетра) эта реакция превращается в положительную под влиянием перехода к активному питанию и образованию соответствующих пищевых условных рефлексов. В последующие дни реакция мальков на свет может индивидуально меняться в зависимости от конкретных условий кормления (В. Ю. Касимов).
В других случаях происходит достройка или изменение врожденного узнавания путем включения новых сенсорных систем. Так, дроздята, как уже упоминалось, в первые дни после вылупления реагируют на сотрясение гнезда выгягиванием шеи и раскрытием клюва. Пространственная ориентация этой реакции осуществляется на основе гравитационной чувствительности, локализованной в органе равновесия во внутреннем ухе, а шея вытягивается вертикально вверх независимо от места расположения источника раздражения. Эта врожденная реакция сохраняется и после прозрения птенцов, в недельном возрасте, хотя теперь уже она осуществляется и на зрительный стимул — появление какого-либо объекта в поле зрения птенца. Только постепенно, по прошествии еще нескольких дней, зрение становится ведущей рецепцией, и в конце концов дроздята научаются четко локализовывать оптический стимул, каковым в норме является родительская особь с кормом, и поворачивать голову в сторону его появления.
У молоди осетровых рыб в течение 1–3 дней после выклева, т. е. до перехода к активному питанию, появляется реотаксисная реакция (ориентация тела по потоку воды), которая в отличие от других рыб сопряжена с тигмотаксисом (ориентация по тактильным раздражениям, обусловленным прикосновениями к посторонним предметам): реотаксис проявляется в основном лишь при контакте малька с субстратом, что препятствует его смыванию течением. Только после появления положительно-фототаксисной реакции тактильная ориентация сменяется оптической (оптомоторной реакцией) и малек становится способным ориентировать свое тело и в толще воды (Д. С. Павлов). Это включение новых сенсорных систем, модификация врожденного узнавания, также происходит в условиях накопления первичного сенсомоторного опыта.
Рассматривая взаимоотношения между врожденным узнаванием (и врожденным поведением вообще) и ранним научением, необходимо постоянно помнить о том, что характер этого взаимоотношения в каждом конкретном случае определяется экологическими особенностями данного вида. Как в количественном, так и в качественном отношении эти взаимоотношения будут существенно различаться, например, у рыб, обитающих в различных условиях освещения, температуры, движения воды и т. д.
Так, у пресноводных рыб, нерестящихся в проточной холодной воде на значительной глубине (сиг), личиночный период короткий, личинки сразу начинают активно плавать, и отрицательный геотаксис, а затем и положительный фототаксис приводят их в поверхностные слои воды. Личинки же рыб, нерестящихся в мелководных, слабопроточных, поросших растительностью водоемах (сазан, щука), долго пребывают на месте, приклеиваясь (выделениями специальных желез) к растениям. Решающее значение в этом смысле имеет врожденное узнавание на основе тигмотаксиса, обеспечивающего тесный контакт животного с субстратом. К поверхности воды эти мальки поднимаются по мере своего развития лишь постепенно, как бы по ступенькам, причем тигмотаксис все больше слабеет, положительный же фототаксис все усиливается. Понятно, что и условия накопления первичного опыта, а тем самым и взаимоотношения между врожденным узнаванием и ранним научением, будут у этих двух групп рыб в корне различными.
Ранний опыт
Облигатное научение
Приведенные выше примеры постнатального научения относятся к упомянутому ранее облигатному научению. Сюда относятся все формы научения, которые в естественных условиях совершенно необходимы для выполнения важнейших жизненных функций, т. е. относящихся к видоспецифическому, инстинктивному поведению. Результаты облигатного научения в одинаковой мере необходимы для выживания всем представителям данного вида, и эта видоспецифичность сближает облигатное научение с врожденными формами поведения. Об этом говорит сайт https://intellect.icu . Облигатное научение и врожденное поведение, в частности врожденное узнавание, теснейшим образом связаны друг с другом в единый комплекс. В этом единстве и находит свое воплощение модификация врожденного поведения ранним опытом. Не только сама способность к облигатному научению, но и конкретные ее проявления генетически фиксированы в такой же степени, как и форма инстинктивного поведения.
Характерным признаком облигатного научения является также то, что оно может осуществляться только на протяжении определенных, так называемых сенсибильных (или чувствительных), периодов онтогенеза. Ни до, ни после этого облигатное научение невозможно. Как еще будет показано, эти периоды начинаются чаще всего очень скоро после появления животного на свет и являются непродолжительными. В отдельных случаях, правда, они приходятся на более поздние сроки и могут обнаруживаться даже у взрослых животных, равно как могут быть и относительно длительными.
В отличие от облигатного научения факультативное научение представляет собой, как нам также уже известно, приобретение индивидуального опыта, который зависит от частных условий жизни особи и не является необходимым для всех представителей данного вида в качестве компонента их инстинктивного поведения. Факультативное научение модифицирует, совершенствует и приспособляет видотипичное, врожденное поведение в соответствии с особыми, частными, преходящими, а зачастую и случайными элементами среды обитания особи, поэтому факультативное научение носит сугубо индивидуальный характер, оно не приурочено к определенным сенсибильным периодам и отличается большой лабильностью и обратимостью. Видоспецифичными являются здесь лишь сама способность к научению и пределы этой способности.
Между облигатным и факультативным научением не всегда возможно провести резкую грань, и в реальном поведении животных, особенно молодых, зачастую обнаруживаются сочетания этих двух категорий приобретения индивидуального опыта (см. ниже). Наиболее четко и часто факультативное научение проявляется в виде разного рода навыков, которые уже рассматривались раньше. Что же касается привыкания, то оно, как было показано, может встречаться и в сфере облигатного научения.
Помимо уже отмеченных сфер поведения облигатное научение важно и для формирования пищевого поведения.
Это проявляется прежде всего в том, что путем облигатного научения детеныши во многих случаях узнают отличительные признаки пищевых объектов. Например, экспериментально доказано, что первоначальный прием пищи служит у змеенышей источником накопления опыта, определяющим последующее распознавание пищевых объектов по их химическим признакам. Детенышам морских свинок предъявляли в течение первых девяти дней после рождения различные съедобные и несъедобные объекты, в результате чего и формировалось предпочтение съедобных объектов. Распознавание несъедобных объектов без такого предварительного контакта оказалось невозможным. Это соответствует тому, что в школе Павлова изучалось на щенках (распознавание вкуса и запаха мяса) и получило название «натуральный условный рефлекс».
Отметим в этой связи, что Г. И. Поляков считает такие рефлексы низшей и наиболее древней категорией условнорефлекторной деятельности, функцией старой коры головного мозга и поэтому особенно характерными для пресмыкающихся, у которых условные рефлексы на неестественные, искусственные стимулы вырабатываются лишь с трудом. У млекопитающих же, по Полякову, натуральные условные рефлексы встречаются, как правило, только в раннем постнатальном онтогенезе, когда новая кора еще не созрела. Именно таким путем, например, котенок научается распознавать писк мыши.
Не только типичные признаки пищевого объекта усваиваются детенышами путем облигатного научения. Важное значение это научение имеет и для формирования пищедобывательных приемов, т. е. двигательных актов, обеспечивающих захват и само потребление пищевых объектов, у хищников — овладевание и поедание добычи. Правда, в своей основе эти движения являются врожденными двигательными координациями и созревают независимо от индивидуального опыта, как и другие инстинктивные движения. Однако их достройка, совершенствование и модификация происходят в конкретных условиях постнатального онтогенеза в результате облигатного научения. Так, белка должна научиться наиболее эффективным образом вскрывать орехи, хотя необходимые для этого движения являются врожденными и уже присущи бельчонку в «готовом виде». Детеныши карликовой мангусты (Helogale undulata) оказываются к моменту выхода из гнезда (в 24-дневном возрасте) уже вполне способными к выполнению движений, необходимых для ловли добычи, хотя заниматься этим им еще не приходилось и они только облизывали и обсасывали убитых животных, которых приносили в гнездо родители. Но каждому детенышу приходится учиться правильно производить укус, мгновенно умерщвляющий добычу, а также учиться обращению с реже встречающимися видами жертв. Характерное для этих животных бросание пищевых объектов с твердой поверхностью (особенно яиц) под туловище, назад, осуществляется первоначально в несовершенном виде и несинхронно. И это действие совершенствуется путем научения.
Необходимо, однако, отметить, что подобная «отработка» инстинктивных компонентов поведения происходит у высших животных главным образом на следующем, игровом этапе постнатального развития поведения. Как будет еще показано, игра имеет в этом отношении исключительно большое значение для окончательного формирования поведенческих актов. У низших животных, у которых нет игровой активности и соответственно отсутствует второй этап постнатального развития поведения, облигатное научение выступает как единственный фактор постнатальной достройки инстинктивного поведения. Особенно это относится, очевидно, к беспозвоночным, хотя у этих животных облигатное научение, как и весь онтогенез поведения, еще совершенно недостаточно изучено. По имеющимся отрывочным данным можно, однако, заключить, что во всяком случае у насекомых облигатное научение может играть существенную роль в формировании поведения. Так, английский этолог В. Г. Торп и его сотрудники установили, что имагинальные, половозрелые формы насекомых ориентируются по запахам, влиянию которых они подвергались на определенных стадиях (сенсибильных периодах) их личиночного развития. Если, например, воздействовать на личинок плодовых мушек (дрозофил) запахом мяты, то они будут впоследствии, превратившись во взрослых насекомых, отыскивать для откладывания яиц субстрат с этим запахом. Наездник Nemeritis canescens паразитирует на личинках моли рода Ephestia, но если личинок наездника вырастить на личинках другого вида (Meliphora), который в естественных условиях не поражается этим наездником, то запах этого вида будет затем предпочитаться взрослыми насекомыми при поиске жертвы. Но этот эффект может быть достигнут лишь в том случае, если контакт с необычным хозяином (личинкой Meliphora) устанавливается в момент выхода личинок наездника из яйца или вскоре после этого. Только в этот краткий период запах нового хозяина оказывается действенным. Следовательно, опять же процесс научения приурочен к четко определенному сенсибильному периоду.
Однако запаховая ориентация насекомых определяется не единственно облигатным научением. Так, в указанном эксперименте у наездников сохранилось избирательное отношение к запаху естественного хозяина (Ephestia) наряду с благоприобретенной повышенной чувствительностью по отношению к запаху нового хозяина. Напрашивается вывод, что положительная хемотаксисная реакция в первом случае определяется врожденным узнаванием, во втором — облигатным научением, которое, как всегда, сочетается с этим инстинктивным компонентом поведения и в нормальных условиях подкрепляет и модифицирует его в соответствии с конкретными условиями развития особи.
Запечатление
Как уже отмечалось, ранний опыт слагается прежде всего из облигатного научения. Факультативное научение, если оно вообще происходит, здесь только дополняет, уточняет и конкретизирует процесс облигатного научения. Преобладание последнего в раннем постнатальном онтогенезе объясняется тем, что в этот период происходит достройка врожденных пусковых механизмов ряда важнейших инстинктивных действии путем включения в них индивидуально приобретаемых компонентов. Именно в этом состоит сущность процесса, получившего название запечатления (Pragung, Imprinting).
Запечатление является важным и характерным компонентом раннего постнатального онтогенеза. Это форма облигатного научения (с обязательным присутствием в качестве важных составных частей элементов факультативного научения), при котором очень быстро фиксируются в памяти отличительные признаки объектов инстинктивных поведенческих актов, поэтому запечатление квалифицируют и как «перцептивное научение», направленное на распознавание «незнакомого» в дополнение к «знакомому» (т. е. к врожденному узнаванию). Как и в других случаях облигатного научения, запечатление совершается лишь в течение определенных сенсибильных периодов, причем без пищевого или иного внешнего подкрепления. Результаты запечатления отличаются исключительной прочностью («необратимостью»).
Объектами запечатления являются родительские особи (выступающие одновременно как носители типичных признаков вида), братья и сестры (детеныши одного помета), будущие половые партнеры. Возможно также запечатление внешних признаков постоянных врагов, так как защитная реакция на эти признаки формируется при сочетании вида врага с предостерегающими криками родителей или, как выше отмечалось в отношении мальков рыб, при виде гибели сородича. При более широком понимании запечатления некоторые авторы относят к нему также формирование реакции на пищевые объекты, а также на характерные признаки места обитания (рождения), что, вероятно, не вполне правомерно, хотя процессы запечатления могут и сопутствовать формированию этих реакций. Здесь мы рассмотрим Запечатление лишь в его «классическом» понимании, а именно как связанное с явлениями общения.
Запечатление изучено преимущественно на зрелорождающихся животных, хотя оно свойственно и незрепорождающимся. Чаще всего запечатление происходит вскоре после рождения, в течение весьма ограниченного сенсибильного периода, нередко при первой же встрече с объектом запечатления. Не все ученые придерживаются одного мнения о сущности этого процесса, и некоторые из них (например, Гесс) склонны даже выделить запечатление в особую, отличную от научения категорию поведения. Однако большинство исследователей все же относят запечатление к научению. Особенно четко запечатление проявляется в реакции следования, изучению которой посвятили свои усилия многие ученые.
Реакция следования
Феномен этой реакции состоит в том, что зрелорождающиеся детеныши уже вскоре после появления на свет неотступно двигаются вслед за родителями (и одновременно друг за другом). Этот феномен общеизвестен по поведению детенышей домашних животных. Приведем несколько примеров реакции следования у диких животных.
Самка гоголя, небольшой утки, устраивающей свое гнездо в дуплах на высоте до 15 метров над землей, непосредственно перед вылуплением птенцов покидает гнездо. Она летит к водоему, откуда возвращается лишь спустя несколько часов, но обратно в дупло она уже не влетает, а начинает летать перед входом в дупло из стороны в сторону, издавая призывные крики. В ответ на это птенцы один за другим подскакивают к входному отверстию, откуда они, посмотрев на летящую перед ними птицу, бросаются вниз на землю. Приземлившись, они тут же поднимаются на ноги и начинают ходить.
Интересно отметить, что к подобным поразительным прыжкам способны и птенцы другого вида уток, которые, как правило, устраивают гнезда в норах, — пеганки. Иногда же эти утки гнездятся в местах, расположенных на высоте до 3–5 метров над землей. В этих случаях вылупившиеся утята прыгают, точнее, сваливаются вниз так же, как гоголята. Еще более «драматичные» прыжки совершают птенцы чистиковых, например гагарки, спрыгивающие с высоких крутых утесов, правда, уже в возрасте 19–20 дней.
Даже стукнувшись о прибрежную полосу суши, они тут же поднимаются на ноги и направляются к воде.
Вернемся, однако, к гоголятам. После того как весь выводок покинул описанным способом гнездо, мать, постепенно снижаясь, садится возле него, а затем, немного походив вокруг утят, направляется к водоему, до которого подчас до двух километров ходьбы. Утята сейчас же движутся вслед за матерью и друг за другом, развивая при этом поразительную скорость, а достигнув воды, они незамедлительно пускаются за матерью вплавь. (Сходным образом ведут себя и птенцы гагарок: мать, пролетев над птенцом, садится неподалеку на воду, и он направляется к ней.)
У млекопитающих реакция следования особенно хорошо выражена у копытных, у которых детеныши рождаются в весьма зрелом состоянии и очень быстро приобретают способность стоять и ходить. В связи с большой ролью, которую играет у этих животных обоняние, запечатление у них происходит не только на оптические и акустические, но и на ольфакторные признаки, т. е. на запахи родителей. Например, верблюжонок уже через 10 минут после рождения делает первые попытки подняться на ноги, а еще через 90 минут он уже может свободно стоять. В течение первых суток формируется и реакция следования за матерью. Вальтер описал формирование реакции следования у детенышей антилоп. У изолированных после рождения от матерей детенышей (в неволе) запечатление может произойти по отношению к проходящему мимо человеку, если детеныш увидит его во время сенситивного периода. Правда, советская исследовательница И. З. Плюснина считает, что привязанность к матери формируется у детенышей копытных (имеются в виду ягнята) не только на основе запечатления. Как мы еще увидим, это справедливо и для других животных, в частности для цыплят.
Среди грызунов реакция следования хорошо выражена, например, у детенышей морских свинок. В отличие от подавляющего большинства других грызунов морские свинки не устраивают гнезда, и детеныши рождаются у них вполне развитыми, зрячими. Вес новорожденного составляет 60–120 граммов. Он обладает уже постоянными зубами, поскольку смена молочных зубов происходит еще в утробе матери.
Описана реакция следования и у детенышей тюленей. Тюленята рождаются очень крупными — их величина достигает почти половины длины тела матери. Еще в ее утробе происходит линька и смена молочных зубов. Уже спустя 30 минут после появления на свет новорожденный (и даже извлеченный оперативным путем из тела матери) тюлененок в состоянии ползать подобно взрослым тюленям, плавать, нырять и вылезать на лед. При удалении матери ее детеныши немедленно двигаются вслед за ней. Время от времени мать стимулирует тюлененка, прикасаясь к нему мордой или с силой шлепая передними ластами по воде, обдавая его брызгами. Именно последнее служит «приглашением» к следованию.
Встречается реакция следования также у некоторых рыб, например у цихлид, о чем еще пойдет речь ниже.
Как видно из приведенных примеров, реакция следования ориентирована на родительскую особь (а также на других детенышей того же выводка). Она обеспечивает выживание детенышей, сплоченность животной семьи, возможность ее охраны со стороны взрослых особей и осуществления ухода за потомством. Без реакции следования родительская особь не могла бы осуществить заботу о потомстве, ибо для ее осуществления необходимо, чтобы детеныши в самые сжатые сроки научились отличать индивидуальные признаки своей матери, от которой нельзя отстать, что и достигается путем запечатления. Таким образом, быстрая конкретизация инстинктивного поведения детенышей на индивидуально опознаваемых объектах (родителях, собратьях) обеспечивает здесь формирование жизненно важных приспособительных реакций в максимально сжатые сроки.
Первые опыты по изучению реакции следования проводились еще в 70-х годах прошлого века Д. Сполдингом, но всестороннее изучение и теоретический анализ процессов запечатления, в том числе и реакции следования, были осуществлены Лоренцем в 30-х годах нашего века. С тех пор эти вопросы изучались многими исследователями, но незаслуженно забыто, что уже в то время опыты по изучению реакции следования ставились в лаборатории Боровского в Москве. В этих опытах перед цыплятами передвигалась черная коробка, и если ее «медленно двигали вдоль стола за привязанную к ней веревку, то большинство цыплят шли за ней… шли даже тогда, когда тем самым удалялись от той части стола, где был рассыпан корм».[47] Вместе с тем цыплята не следовали за белой коробкой и черным платком. Изолированные в момент вылупления, цыплята уже спустя сутки не реагировали на наседку, хотя та относилась к ним как обычно, но вместо этого следовали за рукой экспериментатора. В лаборатории Боровского ставились и другие опыты по этой теме, но затем лабораторное изучение реакции следования было в Советском Союзе возобновлено лишь в 60-х годах (исследования К. Э. Фабри и сотрудников). В ином плане запечатление (на неподвижный объект) изучалось впоследствии в лаборатории А. Д. Слонима.
Еще Сполдинг установил, что при реакции следования объектом запечатления, т. е. объектом видотипичных, инстинктивных действий, может быть не только любой представитель данного вида, но и другое животное или человек и даже неживой, но подвижный объект. Важно лишь, чтобы это был первый передвигающийся объект, увиденный в жизни. Цыплята, у которых была сформирована реакция следования за двигающимся шаром, не станут затем следовать за другим объектом или даже шаром, но иного цвета, а будут избегать его и неотступно двигаться вслед за «своим» шаром.
Как и в других случаях запечатления, реакция следования формируется в сжатые сроки: у кур и уток сенсибильный период начинается в первые же часы после вылупления, достигает оптимума через 10–15 часов, а затем постепенно сходит на нет. В приведенном выше примере выпрыгивание гоголят из дупла происходило именно в оптимальные часы сенсибильного периода, когда со времени вылупления еще не прошло 12 часов. У морских свинок сенсибильный период весьма растянут (с 6-го до 30–40-го дня жизни), но оптимальный срок — 6–7-й день. До 14-го дня «запечатляемость» еще сохраняется на довольно высоком уровне, но затем постепенно сходит на нет (И. З. Плюснина).
Уже упоминалось о том, что запечатление совершается с исключительной быстротой. Следовать за объектом запечатления птенцы начинают зачастую уже после первой встречи с ним, т. е. в эксперименте — после первого же показа. Но если даже и потребуется несколько предъявлений, реакция быстро достигает максимальной интенсивности и в дальнейшем уже остается на этом уровне до тех пор, пока птенцы не подрастут.
Большой интерес представляет вопрос о подкреплении реакции следования. Ведь детеныши следуют за объектом запечатления без всякого вознаграждения, а если искусственно затруднить следование или даже нанести птенцам болевые раздражения, то реакция следования становится только еще более интенсивной (Гесс). Объясняется это тем, что в отличие от факультативного научения облигатное научение не зависит от наличия или отсутствия не только случайных, маловажных компонентов среды, но и таких, которые могли бы служить для подкрепления результатов научения. Результаты облигатного научения настолько важны для жизни особи, что соответствующие реакции должны срочно формироваться при любых обстоятельствах, даже если в данный момент отсутствует возможность их внешнего подкрепления. Правда, при запечатлении имеет, очевидно, место «внутреннее» подкрепление по принципу обратной связи, источником которого являются ощущения от самих производимых движений, т. е. происходит лишь проприоцептивное подкрепление.
Отмеченная выше «необратимость» запечатления проявляется в чрезвычайной прочности реакции следования: практически объект запечатления не может в этом качестве заменяться другим объектом, и «реимпринтинг», т. е. переучивание на другой объект, как правило, возможен лишь в специальных лабораторных условиях и то лишь с большим трудом. Таким образом, чрезвычайно быстро сформировавшееся следование за объектом с определенными отличительными признаками в дальнейшем уже существенно не меняется. Лишь к концу действия запечатления (когда выводок распадается) ослабевает первоначальная интенсивность этой реакции, образовавшаяся без внешнего подкрепления и не нуждающаяся в таковом для своего сохранения. Именно такую характеристику давал процессам запечатления в свое время Лоренц.
Половое запечатление
По-иному проявляется запечатление в сфере размножения. Здесь у многих животных наблюдается так называемое половое запечатление, которое обеспечивает будущее общение с половым партнером. Главная особенность полового запечатления заключается в том, что окончательный результат проявляется лишь с большой отсрочкой, ибо животное учится распознавать отличительные признаки будущего полового партнера еще на раннем этапе постнатального развития. При этом запечатление типичных признаков полового партнера должно совершиться у детеныша в таком виде, в каком они предстанут перед ним уже во взрослом, половозрелом состоянии.
В основном половое запечатление наблюдается у самцов, причем запечатляются у них отличительные признаки их матерей в качестве «образцов» самок своего вида. Следовательно, на врожденное узнавание общих видотипичных признаков здесь путем облигатного научения (полового запечатления) накладывается распознавание видотипичных женских признаков. Так еще в раннем возрасте происходит объективация будущего полового поведения.
В настоящее время факты полового запечатления установлены уже у различных позвоночных, но в первую очередь у птиц. Так, немецким этологом Ф. Шутцем было, в частности, показано, что у самцов диких уток сенсибильный период полового запечатления простирается от 10-го до 100-го дня с момента вылупления, но оптимальный срок ограничен 10–40 днями (в естественных условиях утиная семья распадается на 8–10-й неделе после появления утят). По другим данным этот срок может быть и несколько более длительным. Если на протяжении этого срока птенец мужского пола видел только самку другого вида, он по достижении половозрелости будет «ухаживать» только за самками этого вида, оставляя без внимания уток своего собственного вида. Из гусей, по данным Шутца, половое запечатление лучше всего проявляется у серого, а также и белолобого гуся.
Исследованиями шведского этолога Л. Б. Фэлта было установлено, что и у птенцов диких уток женского пола возможно половое запечатление, если их изолировать от селезней на протяжении первых восьми недель после вылупления. Если им при этом дать возможность видеть самцов-альбиносов или даже людей, то впоследствии нормальное половое поведение уток будет направлено на них, но не на селезней в их обычном брачном наряде. Половое запечатление на человека у представителей этого отряда (гусиных) в свое время было описано и Лоренцем. Известен аналогичный случай и в отношении самца рыжей цапли, выросшего с 10-дневного возраста в домашних условиях и проявившего все формы брачного поведения по отношению к человеку, вырастившему его.
Половое запечатление на альбиносов (у птенцов мужского пола) удалось экспериментально доказать и у зебровой амадины (вьюрка), т. е. незрелорождающейся птицы: птенцы, выращенные родителями-альбиносами, предпочитали впоследствии спариваться с самками-альбиносами, а не с нормально окрашенными особями. Аналогичные результаты были получены и в экспериментах над голубями, когда птенцы белой окраски первоначально выращивались вместе с темноокрашенными взрослыми и наоборот. После изоляции, длящейся с 40-дневного возраста до половозрелости, большинство самцов выбирало самок того цвета, который был свойствен особям, с которыми они в первые недели своей жизни выращивались.
Еще более четкие результаты дают опыты с заменой кладок, при которых птенцы с самого рождения выращиваются приемными родителями. При такой постановке эксперимента у голубей удавалось получить половое запечатление даже в отношении чужого вида. Интересно, что эффект этого запечатления постепенно ослабевает, хотя в какой-то мере еще обнаруживается даже у 5-летних голубей. Таким же способом была установлена возможность полового запечатления на чужой вид у двух вьюрков, легко размножающихся при клеточном содержании (Lonchura striata, Taeniopygia castanotis). И у этих птиц половое запечатление в полной мере проявляется у самцов, которые игнорируют в половозрелом состоянии самок собственного вида, но у самок половое поведение в значительно большей мере определяется врожденными механизмами (К. Иммельманн). Е. В. Лукина наблюдала нечто подобное у канарейки и чижа.
Половое запечатление вообще является весьма сложным процессом и по-разному проявляется у разных видов птиц. Например, оно играет, очевидно, меньшую роль у куриных, как это вытекает из экспериментов П. Гитона, показавшего, что условия выращивания не сказываются на последующем выборе петухами самок. Узнавание отличительных признаков противоположного пола является у этих птиц, очевидно, вполне врожденным.
У млекопитающих половое запечатление изучено главным образом у копытных и грызунов. Имеются, например, указания на то, что скрещивание между ламами викуньей и альпакой возможно лишь в том случае, если новорожденному представителю первого вида дают в качестве кормилицы самку альпака. Немецкие этологи Д. и Х. Мюллер-Шварце показали, что у чернохвостого оленя в половом запечатлении главную роль играют запахи. Группой итальянских исследователей проводились опыты, в которых мыши-родители опрыскивались пахучими веществами. В результате их детеныши, будучи половозрелыми, оказались не в состоянии различать пол сверстников, т. е. не могли найти полового партнера. Аналогичные результаты были получены и у крыс. Опыты с применением «приемных родителей» производились и с морскими свинками. Эффект полового запечатления на чужой вид был выявлен у самцов, отделенных в течение первой недели жизни от своих матерей.
Интересно отметить, что половое запечатление может иметь место и у взрослых особей. Доказано это для некоторых рыб, например меченосца (Xiphohorus maculatus) — известной аквариумной рыбки из отряда карпозубых. Самцы этого вида, как показали шведские этологи А. Ферне и С. Сьеландер, предпочитают самок той окраски, которую им приходилось видеть в течение двух месяцев после наступления половозрелости, но не раньше, как это имеет место в описанных выше случаях полового запечатления.
Подводя итог обзору процессов запечатления, важно подчеркнуть, что общим и наиболее существенным для всех форм запечатления является быстрая постнатальная достройка врожденного поведения, точнее, врожденных пусковых механизмов путем их дополнения индивидуально приобретаемыми компонентами. В результате инстинктивное поведение конкретизируется на определенных, индивидуально опознаваемых объектах, чем и обеспечивается эффективность выполнения инстинктивных действий.
Раннее факультативное научение
Как самостоятельная категория приобретения индивидуального опыта факультативное научение играет в раннем онтогенезе еще значительно меньшую роль, чем на последующих этапах онтогенеза. Это объясняется тем, что оно первоначально лишь дополняет облигатное научение.
Экспериментально факультативный компонент раннего научения можно выявить, воздействуя на детенышей искусственными стимулами, которые или совершенно не встречаются в нормальной среде обитания данного вида, или хотя и могут встречаться, не обладают сколько-нибудь существенной биологической валентностью и не обусловливают определенную инстинктивную реакцию. Кроме того, этот компонент может быть выявлен путем обучения животных действиям, не представленным в их видотипичном поведении.
В качестве примера можно привести эксперимент, в котором крысят в возрасте 5, 7, 9 и 11 дней обучали с помощью электрического раздражения преодолевать коридор, причем ток выключался только при достижении крысятами стенки в конце коридора. При всех прочих оборонительных реакциях (поворотах, попытках забраться на стенку, опрокидывании на бок) действие болевого раздражителя продолжалось. У всех подопытных животных по мере тренировки число таких реакций уменьшалось, но только 7– 11 — дневные крысята научились все быстрее находить правильное направление к стенке в конце коридора и тем самым скорее избавляться от действия тока. Это говорит о том, что до 5-дневного возраста факультативное научение проявляется в еще весьма ограниченных рамках и неспособно помочь животному в выборе адекватного направления действия. Еще позже у крысят появляются факультативные реакции на положительные раздражители. Так, например, приблизительно только с 20-дневного возраста их можно научить нажимать на рычаг за пищевое вознаграждение. Однако, по данным ряда исследователей, оптимальный период раннего постнатального научения начинается у них в месячном возрасте.
У хищных млекопитающих отмечаются аналогичные сроки формирования первых положительных навыков. У щенков было установлено, что прогрессивное развитие способности к формированию навыков в раннем постнатальном онтогенезе непосредственно зависит от развития кратковременной памяти, что в свою очередь связано с развитием коркового торможения (Р. Г. Лозовская и Г. А. Образцова).
У других незрелорождающихся млекопитающих, которые появляются, однако, на свет с уже вполне функционирующими дистантными рецепторами, факультативное научение начинается значительно раньше. Так, по данным советской исследовательницы Н. А. Тих, много лет изучающей психику обезьян в сравнительно-психологическом аспекте, у низших обезьян в условиях искусственного выращивания уже на 3–4-й день после рождения появляются специфические реакции на вид рожка, из которого кормят детеныша, в частности своеобразные движения рук для его обхватывания (после 40–50 кормлений). Экспериментальный условный рефлекс на звук Тих удалось выработать у детеныша гамадрила в возрасте 29 дней.
Необходимо, конечно, учесть, что первые условные рефлексы на тактильные, термические или проприоцептивные раздражители образуются и у других млекопитающих в первые же дни после их рождения. Не исключено, следовательно, что в сфере действия таких раздражителей с самого начала постнатального развития может иметь место не только облигатное, но и факультативное научение даже у животных, рождающихся в крайне незрелом состоянии. Само собой разумеется, что именно так обстоит дело у всех зрелорождающихся животных.
Что касается низших позвоночных, то, как показали некоторые исследования, в частности В. А. Трошихина, экспериментальная выработка стойких условных рефлексов на искусственные раздражители удается уже у мальков рыб. Так, для образования условного пищевого двигательного рефлекса на вспыхивание электрической лампочки у малька карпа достаточно в среднем 4–6 сочетаний, причем положительная условная реакция остается стойкой в течение 100 повторений светового раздражителя без пищевого подкрепления.
Взаимоотношения между компонентами раннего постнатального развития поведения
Все компоненты раннего постнатального развития поведения переплетаются друг с другом, в результате чего это развитие приобретает очень сложный характер. Проанализируем эти связи и взаимозависимости на приведенном выше примере поведения вылупившихся гоголят.
В поведении этих птенцов прежде всего обращает на себя внимание врожденная (инстинктивная) двигательная реакция — подскакивание птенцов к выходу из дупла в ответ на призывные крики матери, а также врожденное узнавание светлого отверстия как цели прыжка. Врожденный характер подскакивания подтверждается работами норвежского исследователя Т. Бьерке, установившего, что для его появления необходим определенный уровень физиологической «зрелости» птенца. Но едва ли имеет здесь место также всецело врожденное узнавание призывного крика, так как еще до вылупления птенцов между ними и материнской особью, вероятно, устанавливается уже знакомый нам взаимный акустический контакт, как это доказано для других зрелорождающихся птиц. Следовательно, нельзя исключить возможность своего рода акустически-кинестетического эмбрионального запечатления голоса матери, т. е. пренатального облигатного научения, как его теперь можно назвать. С другой стороны, совершенно очевидно, что птенцы не могли в период эмбриогенеза тренироваться в целенаправленных прыжках.
Далее существенно, что гоголята реагируют на звуки матери еще до того, как увидят ее: только высунувшись из дупла, они впервые видят ее летящей перед собой. Также врожденная реакция спрыгивания на землю непосредственно не связана с ее присутствием (хотя, возможно, и стимулируется ее видом и голосом), так как подобным же образом ведут себя и гоголята, выращенные в инкубаторе, когда они оказываются на светлом месте (наблюдения Фабри).
Завершается этот первый постэмбриональный этап собственно реакцией следования утят за удаляющейся к водоему матерью (и друг за другом). Эта реакция наступает в ответ на вид двигающейся (в данном случае ходящей) птицы в сочетании с издаваемыми ею звуками, что четко доказано экспериментами, проведенными многочисленными исследователями не только на утятах, но и на цыплятах и птенцах других зрелорождающихся птиц.
Выше уже отмечалось, что конкретное значение запечатления типа реакции следования состоит в индивидуальном узнавании особенно важного компонента среды — родительской особи. Ясно, что это не может осуществляться только врожденным образом, так как распознаваться и запоминаться должны в данном случае не видовые, а именно индивидуальные признаки, т. е. частные отклонения от видотипичной нормы. Объектами врожденного узнавания могут быть лишь общие всем представителям данного вида признаки, а между тем детеныш должен быть в состоянии отличить своего родителя как раз от других особей того же вида. Этому можно только научиться, на это и направлено облигатное научение.
В некоторых случаях, правда, видотипичные признаки, хотя бы частично, тоже фиксируются в памяти детеныша путем запечатления. Особенно, как мы видели, это проявляется при половом запечатлении. В этом состоит одно из важнейших отличий полового запечатления.
Важно отметить, что процессы раннего облигатного научения, прежде всего запечатления, всегда сочетаются с врожденным узнаванием. При этом врожденное узнавание направлено на общие существенные признаки объектов видотипичных действий, путем же облигатного научения эти действия конкретизируются на частном объекте. Так, при реакции следования восприятие детенышем самой подвижности родительской особи (как и любого другого впервые увиденного объекта) является врожденным. Это обусловлено тем, что подвижность представляет собой в данном случае существенный признак объекта запечатления. Индивидуальные же отличительные признаки родительской особи (т. е. объекта видотипичного действия) заучиваются путем запечатления. Следовательно, слияние врожденного поведения (врожденного узнавания) с облигатным научением служит основой дальнейшего развития видотипичного поведения.
Вместе с тем в запечатлении содержатся элементы и факультативного научения. Так, например, птенцы одного и того же выводка могут научиться узнавать родительскую особь по разным ее признакам, и при этом не имеет значения, по каким конкретно признакам ориентируется тот или иной птенец. Облигатным, обязательным здесь является то, что каждый птенец должен вообще научиться узнавать родителя по каким-либо его индивидуальным признакам.
Сочетание врожденных и приобретаемых компонентов сказывается здесь еще и в другом. Как показали эксперименты, проведенные Фабри на цыплятах, первоначальный эффект врожденного узнавания при реакции следования недолговечен и без постоянного упражнения довольно быстро угасает. Однако этот эффект, т. е. следование, можно легко восстановить путем факультативного научения, направленного на объект запечатления. В упомянутых экспериментах объектом запечатления служил для одних цыплят синий шар, для других — желтый цилиндр. После первых же предъявлений цыплята стали следовать за этими объектами не менее интенсивно и продолжительно, чем в обычных условиях за курицей. Но даже наиболее активные подопытные цыплята переставали следовать за объектом запечатления уже через несколько дней, если прекращались ежедневные тренировки. В обычных же условиях, как известно, эта реакция сохраняется в течение ряда недель. Однако и в искусственных условиях эксперимента цыплята тем дольше продолжали следовать за объектом запечатления, чем больше было всего пробежек, т. е. длительность сохранения реакции следования оказалась прямо пропорциональной интенсивности локомоторной активности (частоте пробежек).
В дальнейшем, уже после того, как реакция следования у цыплят совсем исчезла, у них в специальной установке были выработаны навыки следования за бывшим объектом следования — синим шаром или соответственно желтым цилиндром. Следование за этим объектом подкреплялось пищей. Другими словами, следование было заново выработано у цыплят, но путем факультативного научения.
Результаты этой части эксперимента показали, что у цыплят, у которых раньше была сформирована реакция следования путем запечатления, этот навык образовался намного быстрее, чем у цыплят, которые до этого вообще не подвергались запечатлению, т. е. не имели возможности следовать за каким-либо подвижным объектом или же следовали за объектом запечатления другой группы подопытных животных (для цыплят, запечатленных на синий шар, это был желтый цилиндр и наоборот, — рис. 21).
Из этих данных, казалось бы, следует, что врожденное действие (само следование), протекание которого не зависит от побочных внешних факторов и сроки проявления которого генетически фиксированы, угасает без упражнения так же, как это происходит с навыками, а с другой стороны, навык, факультативное научение, может вновь «оживить» эту инстинктивную реакцию. Объясняется это, очевидно, тем, что существует не одна, а две генетически различные, но функционально тождественные формы следования — истинно врожденная и индивидуально приобретаемая, причем вторая форма образуется путем чисто факультативного научения (ведь курица подкрепляет следование своих цыплят кормом и обогревом). Очевидно также, что первая форма со временем все больше перерастает во вторую, иными словами, реакция следования претерпевает качественную перестройку, теряя свою первоначальную врожденную сущность. Одновременно врожденное узнавание переходит в благоприобретаемое узнавание, основанное на индивидуальном опыте.
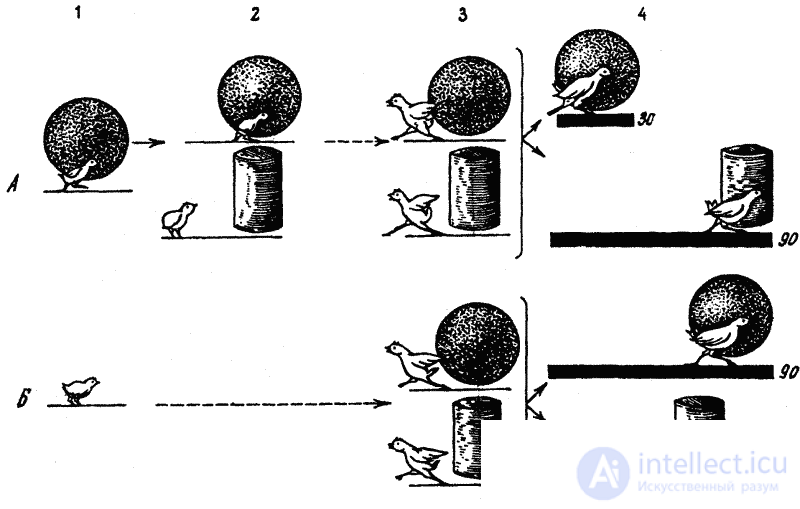
Все это указывает на значение раннего факультативного (а не только облигатного) научения для полноценного проявления инстинктивных элементов поведения. Иными словами, уже на первых этапах постнатального развития врожденное поведение нуждается в участии научения в обеих его формах, которые при этом сливаются с ним. Этим прежде всего и характеризуется сущность всего поведения животных на ранних этапах постнатального онтогенеза.
Раннее формирование общения
Раннее формирование общения у птиц
Ознакомившись в общих чертах с основными компонентами раннего постнатального развития поведения и их взаимоотношениями, рассмотрим теперь специально, как формируется общение между детенышами и родителями и другими особями. Правда, коммуникативные компоненты поведения уже неоднократно встречались нами при ознакомлении с разными аспектами онтогенеза поведения, особенно в процессах запечатления, одним из определяющих факторов которого и является общение.
У птиц, как было уже показано (в частности, в предыдущей главе на кайрах), установление акустических контактов между родительской особью и ее потомством начинается уже на эмбриональной стадии развития птенцов путем пренатального облигатного научения (эмбрионального запечатления). У кайр, например, этот процесс происходит в форме установления акустико-кинестетических связей. В результате этого запечатления птенец подбегает к источнику запечатленного звука. Эта реакция обнаруживается даже у только что вылупившегося птенца: услышав запечатленный звук, он подбегает к родительской особи (в эксперименте к замещающему ее громкоговорителю), издает ответный крик и залезает под нее (или под аппарат). Если же из громкоговорителя после этого раздается другой звук (чужой зов), то птенец немедленно выскакивает из-под него и прячется в укрытии (опыты Чанц). Таким образом, коммуникативный компонент врожденного пускового механизма достраивается в данном случае в ходе эмбрионального запечатления.
Вместе с тем и у родительской особи происходит запечатление индивидуальных особенностей голоса еще не вылупившегося птенца, что обеспечивает индивидуальное опознавание последнего уже к моменту вылупления. Подобное «обратное запечатление» было установлено и у других птиц, а в постнатальном периоде также у млекопитающих и ряда рыб.
Укажем еще на работы Готтлиба, который показал на утятах, что если эмбрион не подвергается в последние дни инкубации соответствующим акустическим воздействиям, то установление звуковых контактов между вылупившимся птенцом и материнской особью будет значительно затруднено. Изучая затем реакцию следования у нескольких видов уток (крякв, пекинских уток, мандаринок), Готтлиб установил, что наиболее активно утята следовали за макетом (чучелом) при видотипичной визуально-акустической стимуляции, хуже — при одной лишь акустической, т. е. без чучела, и еще хуже при одной оптической, т. е. при неозвученном чучеле. Вместе с тем реакция следования значительно ослабевала, когда видотипичные визуальные или акустические компоненты (позывные звуки и внешний вид чучела) подменялись невидотипичными (т. е. принадлежащими другому виду уток). Канадский ученый Р. М. Эванс в опытах над чайками показал, что искажения в реакциях вылупившихся птенцов (предпочитание криков другого вида) могут обусловливаться отсутствием соответствующего эмбрионального опыта, а именно когда эмбрион лишен возможности слышать крики птиц своего вида.
Итак, как правило, вылупившиеся птенцы реагируют четко избирательно на видотипичные крики, т. е. предпочитают ориентироваться по «натуральным» акустическим раздражителям с большой биологической валентностью. Эти реакции основаны на врожденном узнавании (и предпочтении) видотипичных компонентов коммуникации, которое дополняется и уточняется облигатным выучиванием индивидуальных компонентов общения в процессе пре- и особенно постнатального запечатления. Следовательно, при постнатальном формировании акустического общения между птенцами и их родителями возможность индивидуального (не врожденного!) опознавания особей обеспечивается ранним облигатным научением. Принадлежность же последних к одному и тому же виду устанавливается птенцами на основе врожденного узнавания.
На основе данных, полученных рядом ученых, сейчас можно, очевидно, считать установленным, что и другие функции пения птиц формируются в большой степени благодаря постнатальному облигатному научению. Например, у овсянки песня молодого самца приобретает сексуальную, а затем и территориальную функцию только после того, как он научается петь «полную» песню, т. е. модифицировать ювенильную «подпесню», придать ей специфическую структуру и видоизменить ритм.
Большую роль играет при этом подражание, на что, в частности, еще в середине 50-х годов указывал Торп, отмечая, что птицы учатся непосредственно по звуковым сигналам других птиц.
Выше уже рассматривались явления подражания и говорилось о необходимости выделить как отдельные категории облигатное и факультативное имитационное научение. Теперь должно быть ясно, что облигатное подражание, будучи особым проявлением облигатного научения, характерно прежде всего для раннего постнатального периода онтогенеза. В области звукового общения это можно хорошо проиллюстрировать примерами из многочисленных полевых экспериментов, осуществленных советскими орнитологами К.А. и К. К. Вилкс путем перемещения яиц и птенцов в гнезда других видов (еще раньше подобные эксперименты проводились Промптовым). Изучив поведение приблизительно 4000 пересаженных воробьиных птиц (преимущественно мухоловок-пеструшек), эти исследователи установили, что птенцы в исключительно широком диапазоне перенимают песни «приемных родителей»: более 80 % самцов, выросших в гнездах других видов, имитировали песню вида, к которому принадлежала особь, воспитавшая птенца (горихвостки, пеночки-трещотки, большой синицы). Вместе с тем при имитации чужой песни воспроизводились и элементы песни собственного вида. Понятно, что в нормальных условиях акустическое поведение птенца формируется путем облигатного имитационного научения звукам, издаваемым кормящей птицей того же вида.
Примечательно, что перенимание чужих звуков происходит в относительно короткий период начиная с вылета птенца из гнезда. Это доказывается тем, что чужие звуки имитировали и птицы, пересаженные незадолго перед вылетом, но если, наоборот, птенец воспитывался с самого начала в чужом гнезде, но перед вылетом пересаживался обратно к своему виду, чужое воспитание не сказывалось на акустических реакциях такой птицы и ее пение оставалось видотипичным. Таким образом, здесь определенно можно говорить о наличии сенсибильного периода, что опять-таки доказывает, что мы имеем в этом случае дело с облигатным научением, точнее, облигатным имитационным научением.
Раннее формирование общения млекопитающих
У млекопитающих взаимное запечатление индивидуальных отличительных признаков родителей и детенышей и установление контактов между ними происходят в разные сроки после появления детеныша на свет — в зависимости от степени зрелорождения, но чаще всего в первые часы жизни новорожденного. Детеныш одногорбого верблюда, например, издает первые звуки еще во время самих родов, а уже час спустя он в состоянии воспроизвести почти все звуки, свойственные его виду, поэтому уже с момента рождения начинается интенсивный акустический контакт между ним и его матерью.
Американским исследователем онтогенеза поведения животных Дж. П. Скоттом и рядом других ученых было доказано, что у собак и других видов псовых существует сенсибильный период, в течение которого только и возможно такое установление контактов с материнской особью и остальными щенками данного выводка. Очевидно, при этом процесс запечатления имеет комплексный характер, поскольку включает в себя и половое запечатление. На это указывает и то обстоятельство, что общение между матерью и щенками, равно как между последними и другими представителями того же вида, может быть по форме идентично половому общению (М. В. Фокс). Так же, по свидетельству ряда ученых, обстоит дело и у обезьян.
Большое значение для формирования общения имеет положительная тигмотаксисная реакция детенышей, выражающаяся в активном установлении физического контакта с поверхностью матери и собратьев, в прижимании к ним. Об этом свидетельствуют, в частности, эксперименты, проведенные на детенышах обезьян Г. Харлоу и его сотрудниками, применявшими разного рода макеты материнских особей (проволочные, тряпочные, обогревамые, холодные и пр.). Было показано, что детеныши предпочитают запечатленный холодный макет даже обогреваемому. Сходная тигмотаксисная реакция, как показали английский этолог В. Слэкин и другие исследователи, присуща также цыплятам и другим птенцам.
Скотт пришел к выводу, что у млекопитающих в раннем постнатальном онтогенезе существуют сенсибильные периоды трех типов: для процессов научения, стимуляции физиологических процессов и формирования общения. Исследования многих ученых показали, что если в течение сенсибильного периода последнего типа детеныш не имеет возможности установить связи с особями своего вида, он впоследствии может оказаться совершенно неспособным к общению с себе подобными и во всяком случае будет испытывать большие затруднения в коммуникативном, а также воспроизводительном поведении. То, что формирование первичных коммуникативных связей совершается путем запечатления, подтверждается и тем, что оно, как доказано рядом экспериментаторов, не зависит от пищевого или иного внешнего подкрепления.
Не менее четко выражено у млекопитающих и «обратное запечатление» — запечатление индивидуальных признаков детенышей у родителей. Так, например, овцы, козы и другие копытные научаются распознавать эти признаки непосредственно после рождения их детенышей и после этого уже не подпускают к соскам чужих. Ведущее значение имеет здесь, вероятно, ольфакторное опознавание, т. е. запоминание индивидуального запаха потомства, хотя и оптические признаки могут играть существенную роль. У детеныша, как показали в эксперименте с чернохвостыми оленями Д. и Х. Мюллер-Шварце, также происходит запечатление индивидуального запаха матери или, в эксперименте, заменяющей модели, причем в отношении последней вполне осуществляются акты общения, проявляемые в норме по отношению к подлинной матери.
Формирование общения у низших позвоночных
Отмеченные закономерности не имеют конечно, значения всеобщих правил для всех позвоночных, а тем более беспозвоночных. Даже среди птиц и млекопитающих обнаруживаются многочисленные отклонения и исключения, обусловленные экологическими факторами, особенностями образа жизни. С другой стороны, эти закономерности проявляются и у многих рыб. Так, например, мальки цихлид также научаются опознавать своих родителей путем запечатления и неотступно следуют за ними, но только тогда, когда те находятся в брачном наряде. Таким образом, облигатное научение здесь более крепко связано с врожденным узнаванием и более ограничено непосредственным действием ключевых раздражителей (в данном случае светло-темные контрасты, зеленые и синие цвета, движение), чем это наблюдается у высших позвоночных. Доминирующую роль здесь играют врожденные пусковые механизмы, индивидуальное же распознавание играет второстепенную роль. Средствами общения являются при этом сами движения, выполняемые родителями и мальками и имеющие генетически фиксированное сигнальное значение.
Что же касается «обратного запечатления», то оно у цихлид не связано с индивидуальным опознаванием потомства родительскими особями. Последние узнают мальков лишь как таковых, но неспособны отличить свой выводок от чужого, как это показали опыты с перекрестным воспитанием молоди путем замены родителей в семейных группах. При этом более крупная молодь поедается. Если подрастающих мальков повторно заменить более молодыми (т. е. более мелкими), то уход за потомством со стороны родителей можно растянуть на срок до 9 месяцев. На этой фазе преобладает оптическое общение, на личиночной — химическое (опыты Д. Наукса и Дж. Барлоу).
У низших позвоночных, в частности у рыб, присутствие особей того же вида не является столь необходимым для формирования форм общения, как у высших животных. В этом также проявляется более жесткое врожденное «программирование» раннего постнатального поведения рыб, чем у последних. Сходное положение наблюдается, очевидно, и у пресмыкающихся. Так, у ящериц-анолисов важный компонент общения — кивание головой с одновременным вытягиванием ног — наблюдается уже у новорожденных (через 5 минут после вылупления), причем и у изолированных животных, лишенных возможности видеть сородичей. Таким образом, здесь налицо подлинные врожденные двигательные координации, жестко регламентирующие коммуникативное поведение без индивидуального опознавания сородичей.
Познавательные аспекты раннего постнатального поведения
Исследовательское поведение
Постоянной составной частью любого поведенческого акта является исследовательское поведение, проявляющееся на разных онто- и филогенетических уровнях в весьма различных формах. Диапазон этих проявлений простирается от элементарных ориентировочных реакций до исследовательской деятельности высших млекопитающих.
В широком смысле ориентировочное, поисковое поведение является начальной фазой любого инстинктивного действия, и оно направлено на поиск тех агентов среды, которые являются объектами врожденного узнавания. Иначе говоря, в инстинктивных поведенческих актах ориентировочно-исследовательская активность служит поиску и обнаружению ключевых раздражителей и направляет на них деятельность животного. Об этом уже говорилось при изложении этологической концепции инстинктивного поведения.
Элементарные ориентировочные реакции проявляются очень рано даже у незрелорождающихся детенышей. Так, у щенят и лисят уже в первый-второй день жизни наблюдаются поисковые маятникообразные движения головой, которые прекращаются после нахождения соска матери. Сходные поисковые движения, повороты головы в стороны и тыканье концом морды в окружающие животное объекты производятся в первые же сутки жизни также котятами и многими другими детенышами хищных млекопитающих. Детеныши обезьян уже в первые дни жизни двигают ушными раковинами и т. д.
В дальнейшем появляются такие ориентировочные реакции, как обнюхивание, прислушивание, присматривание и т. п. У щенят и лисят такое обследование окружающего пространства начинается приблизительное конце второй недели жизни. У детенышей низших обезьян начиная с 14–16-го дня жизни появляются двигательные реакции в ответ на «индифферентные» звуковые и зрительные раздражители, которые потом (приблизительно в 1,5–2-месячном возрасте) вырастают в подлинные исследовательские действия: животное живо реагирует на изменения в окружающей среде (появление нового раздражителя, нового объекта) поворотами головы, особенно глаз, движениями ушных раковин, а также приближением к новому, незнакомому объекту. Так появляется «любопытство», столь характерное прежде всего для приматов, а также для других высших позвоночных.
Развитие исследовательского поведения в большой мере определяется условиями, в которых растет детеныш, особенно возможностями общения с матерью и другими сородичами. В экспериментах с длительным (до 30 недель) изолированным выращиванием детенышей макаков-резусов (опыты американских ученых Ч. М. Байзингера, Е. М. Брандта и Дж. Мичелля) было установлено, что изолированные детеныши больше рассматривали окружающие их предметы, человека, наблюдавшего за ними, и самих себя, чем нормально живущие со своими матерями. Первые также больше манипулировали предметами и больше трогали самих себя, вторые больше, быстрее и ловчее передвигались по клетке. Чехословацкие исследователи Б. Семигиновский, З. Халупка и И. Мысливечек, ставя опыты над крысятами, нашли, что возможность усиленного общения с сородичами в течение первых 6 недель жизни несколько тормозит развитие исследовательской активности, а обогащенная среда, наоборот, стимулирует его.
Уже на начальных стадиях постнатального онтогенеза наряду с реакциями на появление новых раздражителей имеет место активный поиск последних (маятникообразные движения головой у щенков), который особенно характерен для исследовательского поведения. Лежащие в основе этого поведения ориентировочные рефлексы отличаются, как показали исследования Е. Н. Соколова, значительной сложностью их физиологических механизмов. По Соколову, ориентировочные реакции возникают в результате несовпадения воспринимаемых новых сигналов с формирующимися в головном мозгу «нервными моделями стимула». При этом происходит оценка нового стимула, побуждающая животное к выполнению определенных действий или к их изменению. Наряду с этим ориентировочная деятельность играет также существенную роль в обеспечении необходимого уровня физиологической активности центральной нервной системы.
Нас же в данном случае особенно интересует познавательное значение ориентировочно-исследовательской деятельности, ее роль в приобретении новой информации о компонентах окружающей среды, в обогащении индивидуального опыта животного.
Облигатное научение и ориентация
Рассмотрим сначала некоторые процессы, связанные с начальной ориентацией у детеныша. У всех животных здесь первостепенное значение имеют таксисы, которые, как уже было показано, у высших животных дополняются и обогащаются элементами научения.
Это научение носит прежде всего характер облигатного научения, ибо, как только что отмечалось, оно входит как обязательный компонент в любой инстинктивный акт. Однако каждое животное вынуждено самостоятельно выучивать значимые лишь для него ориентиры, которые будут для разных особей того же вида весьма различными. Отличительные признаки этих ориентиров являются сами по себе случайными, несущественными, и только индивидуальное запоминание их в результате факультативного научения придает им опознавательное значение. Уже говорилось о том, что их выбор животным определяется в большой степени индивидуальными особенностями последнего и разные особи могут предпочитать разные компоненты среды в качестве ориентиров.
Таким образом, в ориентировочном поведении детеныша всегда присутствуют элементы облигатного и факультативного научения, однако соотношения между этими двумя компонентами, их удельный вес могут быть различными в зависимости от того, в какой функциональной сфере совершается ориентация. Когда детеныш учится распознавать «живые ориентиры», особенно внешние признаки своих сородичей, т. е. когда ориентация сочетается с общением, явно преобладают моменты облигатного научения. Это вполне понятно, ибо когда животное ориентируется по подаваемым другими животными оптическим, акустическим, ольфакторным и иным сигналам, которые входят в репертуар общения, то решающее значение имеют наследственно фиксированные, врожденные элементы поведения. При формировании коммуникативного поведения в онтогенезе первостепенное значение имеет поэтому срочная постнатальная достройка соответствующих врожденных пусковых механизмов, что, как мы уже знаем, является главной характерной чертой запечатления. Ограничимся здесь отсылкой и к другим аспектам и примерам облигатного научения с ориентационным значением (например, привыкание к «биологически нейтральным» раздражителям), о которых речь шла выше.
Раннее факультативное научение и ориентация
Уже в раннем ориентировочном поведении заметно сказываются индивидуальные особенности животного. В большой степени индивидуальные различия в поведении зависят от частоты и характера осуществленных с момента рождения сенсорных действий. Это достаточно четко проявляется при выращивании детенышей в условиях, когда им приходится постоянно видеть определенные фигуры. Как показали экспериментальные исследования, животные, выросшие в таком окружении, впоследствии легче ориентируются по таким фигурам. Правда, согласно экспериментам американского ученого Р. М. Осволта, существенное значение имеет при этом степень трудности различения таких фигур. В этих экспериментах крысята еще до прозрения помещались в клетки, на стенах которых попарно помещались фигуры: для одних это были горизонтальные и вертикальные полосы, для других — дуги, для третьих — треугольники и круги. Заранее было известно, что последнее сочетание является наиболее трудным для различения этими животными, полосы же — наиболее легким. В таком окружении крысята пребывали до 50-дневного возраста. Контрольные животные выращивались в клетках без фигур (с белыми поверхностями). Когда затем перед крысятами была поставлена задача — произвести выбор между знакомыми фигурами (одна из которых подкреплялась пищей, другая — нет), то оказалось, что ранний сенсорный опыт облегчал решение задачи только тогда, когда животным приходилось ориентироваться по трудноразличимым фигурам (треугольник и круг). Выращивание же с легче различимыми фигурами не давало такого эффекта: соответственные подопытные животные ориентировались по полосам и дугам не лучше контрольных. При этом Осволт отмечает, что влияние «преэкспозиции» стимулов явно приурочено к определенному раннему периоду, что указывает на участие в данном процессе наряду с факультативными также облигатных компонентов латентного научения.
Итак, ранняя факультативная неподкрепляемая визуальная тренировка способствует ориентации при дальнейшем подкрепляемом факультативном научении (заучивании ориентиров), особенно тогда, когда эта тренировка была сопряжена с трудностями и если имели место более сложные сенсорные действия.
В целом приведенные факты ранней пространственной ориентации показывают, как облигатное научение (благо приобретаемая ориентация по биологически валентным элементам среды) сочетается с факультативным научением, дополняется и конкретизируется им (запоминание особенностей местности путем сенсомоторной тренировки и заучивание конкретных ориентиров). При пространственной ориентации описанного типа на первый план выступает факультативный компонент. Иное соотношение наблюдается, как мы видели, в тех случаях, когда животное ориентируется не по признакам местности, а по признакам сородичей или других животных.
Раннее манипулирование
Исключительно большое значение имеет для приобретения и обогащения индивидуального опыта, как и для всей познавательной деятельности животного, манипулирование. Под манипулированием, манипуляционной активностью мы понимаем активное обращение с различными предметами при преимущественном участии передних, реже — задних конечностей, а также других эффекторов: челюстного аппарата, хобота (у слона), хватательного хвоста (у широконосых обезьян), щупалец (у головоногих моллюсков), клешней (у раков) и т. д. Манипулирование проявляется прежде всего в пищедобывательной и гнездостроительной активности животных. Вообще же, за исключением некоторых ориентировочных, сигнальных и защитных движений, при которых меняется лишь внешний вид животного, но само оно остается на месте, все движения, направленные на внешнюю среду, т. е. составляющие поведение, подразделяются на локомоторные (у большинства высших животных — опорно-локомоторные) и манипуляционные.
У высших животных, как еще будет показано, манипулирование выступает как ведущий фактор развития сенсомоторных функций. Это обусловлено тем, что именно в ходе манипулирования животное вступает в наиболее активный контакт с предметными компонентами среды и получает наилучшие возможности для ознакомления с ними, а также для многообразного воздействия на них.
Манипулирование появляется в различные сроки и проявляется по-разному у разных животных. Особенно велики эти различия у зрело- и незрелорождающихся видов. Но в каждом случае, выполняя разнообразные действия с различными предметами (биологически значимыми и «нейтральными»), детеныши получают комплексную информацию об объекте манипулирования, его свойствах, особенно о его физической структуре. Одновременно происходит развитие и совершенствование (путем тренировки) эффекторносенсорных систем животного. Все это делает манипуляционную активность высшей формой ориентировочно-исследовательской деятельности животных.
В полной мере манипулирование развертывается в игровом периоде онтогенеза. В рассматриваемом здесь периоде манипуляционная активность появляется первоначально лишь в самых простых формах, особенно у незрелорождающихся животных. Так, например, детеныши хищных млекопитающих в первые дни или даже недели, а именно до начала функционирования дистантных рецепторов, т. е. до открывания глаз и ушных проходов, в основном спят, а в промежутках преимущественно ползают в поисках соска и сосут.
По данным советского этолога Н. Н. Мешковой, первые движения детенышей носят характер простейшего манипулирования: детеныши трогают лапой родителей и своих собратьев, хватают их ртом в разных местах тела, нажимают на них мордочкой. Все эти действия еще весьма нечеткие и непродолжительные. У лисенка, по исследованиям того же автора, до 12-дневного возраста (до прозрения) манипуляционная (нелокомоторная) активность развивается следующим образом.
Первые манипуляционные движения обнаруживаются уже у новорожденного, но в течение первых двух часов после рождения они существуют лишь в двух формах: 1) прикасания к объекту передним концом головы (конкретно это проявляется в поиске соска) и 2) хватания объекта губами (захват соска и прилегающих участков кожи ртом). Передние конечности в этих действиях участия не принимают и отведены в стороны.
Затем в течение первых двух суток с момента рождения к этим двум формам добавляются еще пять, которые на этом этапе также связаны только с сосанием и сопутствующими ему действиями (расталкивание других детенышей при движении к соску, поиск соска, отгребание шерсти на животе матери, придавливание или ритмичное толкание лапами живота рядом с соском во время сосания). Эти манипуляции характеризуются прежде всего тем, что детеныш начинает действовать передними лапами и появляются боковые движения головы (в частности, при расталкивании собратьев). Передние лапы прикасаются к объекту (телу матери) и передвигаются по нему одновременно с головой или же самостоятельно, без ее участия.
Последнее имеет место в тех случаях, когда лисенок разгребает одними передними лапами шерсть на животе матери или придавливает его сразу обеими лапами.
Далее, вплоть до прозрения, положение существенно не меняется, появляется лишь еще одна форма манипулирования — захватывание объекта с поочередным придавливанием его обеими передними конечностями. Это имеет место тогда, когда лисенок сосет и при этом ритмично нажимает на живот рядом с соском то одной, то другой лапой.
Итак, к моменту открывания глаз лисенок обладает лишь восемью формами манипулирования, которые к тому же выполняются преимущественно головой. Передние конечности играют в основном лишь вспомогательную роль и способны лишь к самым элементарным самостоятельным движениям. Действия, выполняемые лишь одной передней конечностью, на этом этапе онтогенеза еще полностью отсутствуют. Все это свидетельствует о том, что манипуляционные функции головы, в частности челюстного аппарата, опережают в своем развитии формирование функциональной системы передних конечностей, которые на первых порах еще недостаточно сильны для самостоятельного манипулирования предметами (из восьми форм лишь одна выполняется только конечностями). Здесь, как и в дальнейшем ходе онтогенеза, отчетливо проявляются уже отмеченные раньше реципрокные отношения между этими двумя эффекторными системами.
Познавательное значение раннего манипулирования
Как видно из приведенного примера, активность незрелорождающегося детеныша млекопитающего направлена на этом, доигровом, периоде онтогенеза практически только на один объект — тело матери. Лишь попутно начинают появляться действия, направленные на собратьев, и то лишь для устранения помех на пути к соску. Подлинно активного контактирования между детенышами нет (устанавливается в основном пассивный контакт во время совместного отдыха и сна). Активные действия, направленные на другие, особенно «биологически нейтральные», объекты, еще отсутствуют полностью. Это обстоятельство резко сужает познавательное значение манипуляционной активности на раннем этапе онтогенеза, так как детеныш получает лишь ограниченную и однообразную информацию.
Совершенно иная картина наблюдается, естественно, у зрелорождающихся животных, которым раннее развитие эффекторных и сенсорных способностей позволяет уже в первые дни после появления на свет вступать в многообразные отношения с компонентами среды. Этап раннего формирования манипуляционной активности и связанного с этим постепенного созревания познавательных способностей у этих животных выпадает. О биологическом значении этой специфической особенности зрелорождения уже говорилось в начале главы в связи со взглядами Л. А. Орбели.
Несмотря на первоначальную ограниченность познавательной функции манипуляционной активности у незрелорождающихся животных, мы имеем здесь дело с началом процесса, который в ходе онтогенеза приобретает все большее значение для психического развития особи (см. следующую главу). В этой связи интересно отметить, что зрелорождающиеся животные (птицы и млекопитающие) вообще сравнительно мало манипулируют, во всяком случае в ходе дальнейшего онтогенеза этот компонент их поведения, как и другие, сравнительно мало прогрессирует.
Особенно четко значение манипулирования выступает при сравнении его с запечатлением. И в этом и в другом случае мы имеем внешне не подкрепляемую деятельность, причем генетически фиксированную, врожденную (все животные одного вида манипулируют в целом одинаковым образом). И в том и в другом случае эта деятельность специально направлена на распознавание элементов окружающего мира, на различение знакомого и незнакомого (при запечатлении — «своя» мать и «не свои» особи), на формирование индивидуального опыта. В обоих случаях имеет место упражнение и совершенствование эффекторносенсорных способностей, «моторное» и «сенсорное» научение: животные учатся адекватно употреблять свои органы движения и чувств, причем научение это совершается комплексно.
Но существенная разница между запечатлением и манипулированием состоит в том, что запечатление строго фиксировано тем, что оно направлено лишь на объекты видотипичных, инстинктивных действий, в то время как манипулирование не ограничено никакими рамками: объектом манипулирования может быть любой объект вообще. Более того, особая роль манипулирования в развитии психики и заключается в том, что она лишь первоначально в начале постнатального онтогенеза, направлена исключительно на биологически высоко валентные объекты, т. е. объекты инстинктивных действий. Полностью же и в наиболее сложных формах манипуляционная активность развертывается тогда, когда предметная деятельность подрастающего животного распространяется и на «биологически нейтральные» объекты. Это происходит, когда молодое животное начинает играть. Именно тогда начинает сказываться ведущее познавательное значение манипуляционной активности, ее роль как важнейшего фактора «латентного научения», накопления индивидуального опыта «впрок».
Если я не полностью рассказал про развитие психической деятельности в раннем постнатальном периоде? Напиши в комментариях Надеюсь, что теперь ты понял что такое развитие психической деятельности в раннем постнатальном периоде и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Сравнительная психология и Зоопсихология
Ответы на вопросы для самопроверки пишите в комментариях, мы проверим, или же задавайте свой вопрос по данной теме.

Комментарии