Лекция Тесты
Привет, сегодня поговорим про психофизиология восприятия, обещаю рассказать все что знаю. Для того чтобы лучше понимать что такое психофизиология восприятия, сукцессивное восприятие, симультанное восприятие, потенциал действия, дирекционно-чувствительные клетки, эндогенный, экзогенный, симультанный, сукцессивный, поля коры , настоятельно рекомендую прочитать все из категории Психофизиология. Кликните на вариант (или варианты ответов), если он правильный - то будет подсвечен зеленым цветом и вам будет зачислено пару монеток, а если неверный - то красным и будет снята монетка. Удачи в прохождении онлайн теста!
психофизиология восприятия — направление теоретической психофизиологии, занимающееся изучением физиологических механизмов восприятия .
еловек воспринимает окружающий мир при помощи специализированных сенсорных систем — анализаторов. Восприятие — это процесс и результат формирования субъективного образа предмета или явления, действующего на анализатор.
Классификация анализаторов осуществляется на основе местоположения рецепторов. Рецепторы — это специальные чувствительные нервные образования, воспринимающие раздражения из внешней или внутренней среды и перерабатывающем их в нервные сигналы. По локализации рецепторов анализаторы делятся на экстероцептивные и интероцептивные. В первом случае источники стимуляции находятся вне организма, во втором — информация поступает от внутренних систем и органов. Извне на организм действуют зрительные, слуховые, тактильные, вкусовые, обонятельные стимулы, а также сила тяготения (табл. 5.1).
Таблица 5.1.Экстероцептивные анализаторы и их характеристики
| Модальность | Локализация рецепторов | Тип рецепторов | Воспринимаемое качество |
|---|---|---|---|
| Зрение | Сетчатка | Палочки Колбочки |
Освещенность Контрастность Движение Размеры Цвет |
| Слух | Улитка | Волосковые клетки | Высота Сила звука Тембр Локализация звука |
| Равновесие | Вестибулярный орган | Макулярные клетки | Вращение Сила тяжести |
| Осязание | Кожа | Окончания Руффини Диски Меркеля Тельца Пачини |
Тепло Давление Вибрация |
| Вкус | Язык | Вкусовые сосочки на кончике языка Вкусовые сосочки у основания языка |
Сладкий и кислый вкус Горький и соленый вкус |
| Обоняние | Обонятельный эпителий в носу | Обонятельные рецепторы | Цветочный Фруктовый Мускусный Пикантный запах |
Симультанное и сукцессивное восприятие . Все виды восприятия несут информацию о времени, т.е. о том моменте, когда появился стимул и как долго он действовал. Другими словами, восприятие — это процесс, началом которого служит момент действия стимула на рецептор. Завершением восприятия является образ стимула — объекта и его опознание (идентификация). Длительность одного акта восприятия может быть очень короткой, почти мгновенной, особенно, когда объект восприятия хорошо знаком. В таком случае говорят об одномоментном (симультанном) восприятии. Если человек сталкивается с неизвестным стимулом, длительность восприятия может существенно увеличиваться. Требуется время, чтобы провести детальный сенсорный анализ, выдвинуть и проверить несколько гипотез по поводу действующего стимула, и лишь после этого принять решение о том, что же представляет собой воспринимаемый стимул. В этом случае говорят о последовательной обработке информации и сукцессивном восприятии.
Интенсивное изучение нейрофизиологических механизмов восприятия стало возможным в связи с возникновением методов регистрации микро- и макропотенциалов мозга, т.е. активности отдельных нейронов и суммарной биоэлектрической активности мозга. Возможно поэтому исследования механизмов восприятия многочисленны и включают несколько уровней анализа: от единичного нейрона до целого мозга, причем каждому уровню соответствует свой вариант анализа перцептивного процесса. Однако независимо от того, на каком уровне изучаются процессы восприятия, одно из главных мест занимает проблема кодирования. При этом основной вопрос заключается в том, каким образом происходит прием и преобразование сенсорных стимулов и в каком виде отражается воспринятый и преобразованный стимул в ЦНС человека.
Учение Мюллера. Проблема преобразования информации в нервной системе привлекала внимание исследователей очень давно. Первые идеи в этой области были представлены еще в середине прошлого века учением Мюллера о специфической энергии органов чувств. Суть его состояла в том, что чувствительность к раздражению зависит не от воздействующего раздражителя, а от свойств возбуждаемых нервов. Например, зрительный нерв передает ощущение света, даже если его раздражать механическим путем (ударом по глазу). Мюллер и его последователи полагали, что каждое ощущение возникает при разрядах специфических нейронов мозга, имеющих собственные "линии" связи с периферическими органами. Различные комбинации этих элементарных ощущений должны были создавать более сложные виды восприятия. Разумеется, эти представления в основном имеют исторический интерес.
Принципы "Меченой линии". В настоящее время физиология сенсорных систем очень продвинутая (по сравнению с другими разделами) область нейробиологии, тем не менее основная проблема, по сути, не изменилась. Каким образом импульсация, идущая от специализированных рецепторов органов чувств, передает информацию разных типов? Трудность усугубляется тем, что, хотя рецепторы модально специализированы и чувствительны к определенному типу стимуляции (звуку, свету, давлению и т.д.), нервы, по которым "бегут" импульсы, в основном одинаковы, и сами импульсы, распространяющиеся от этих рецепторов в головному мозге, имеют постоянные характеристики. (Хорошо известно, что нервный импульс —
потенциал действия — генерируется нейроном по принципу "все или ничего").
Наиболее простой ответ предполагает, что мозг узнает о типе воздействующего стимула на основании того, в какой конечный пункт назначения в коре больших полушарий приходит нервная импульсация. Так, потенциалы действия, поступающие в зрительные области коры, несут информацию о зрительных стимулах, а сходные импульсы, поступающие в слуховые зоны, — о звуках и т.д. В наиболее полном виде эти представления воплотились в принципе "меченой линии", в соответствии с которым допускается прямая морфологическая связь и соответственно передача информации от рецептора к определенному центральному нейрону, который отвечает за определение качества стимула.
Однако, каким образом мозг различает разные качества каждого из стимулов в пределах одной модальности, т.е. как мозг дифференцирует разные зрительные или разные звуковые раздражители? Такие тонкие различения осуществляются на основе особых форм организации импульсной активности нейронов, которые получили название кодов.
Коды как средства передачи информации. Кодирование информации в нервной системе — это преобразование специфической энергии стимулов (света, звука, давления и др.) в универсальные коды нейтронной активности, на основе которых мозг осуществляет весь процесс обработки информации. Таким образом, коды — это особые формы организации импульсной активности нейронов, которые несут информацию о качественных и количественных характеристиках действующего на организм стимула.
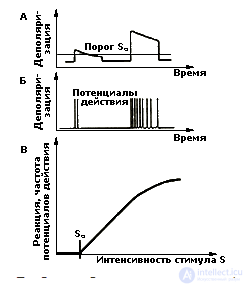
Проблема образования кодов и их функционирования в ЦНС и составляет в настоящее время центральное ядро проблемы представления и преобразования информации в организме человека и животных.
С точки зрения одного из известных специалистов в области сенсорного кодирования Дж. Сомьена (1975) наиболее распространена в сенсорных системах передача информации с помощью частоты разрядов нейронов. Возможны и другие варианты нейронных кодов: плотность импульсного потока, интервалы между импульсами, особенности организации импульсов в "пачке" (группе импульсов) — периодичность пачек, длительность, число импульсов в пачке и т.д. Существует немало данных, подтверждающих, что перечисленные характеристики нейронной активности меняются закономерным образом при изменении параметров стимула. Однако проблема кодирования не сводится только к анализу разных вариантов импульсной активности нейронов. Она намного шире и требует более углубленного анализа (см. Хрестомат. 5.1).
В настоящее время существуют вполне определенные представления о конкретных нейронных механизмах, осуществляющих сенсорный анализ и построение сенсорной модели внешней среды. Они связаны с так называемой концепцией детекторного кодирования.
Детекторная концепция. Главным понятием в детекторной концепции кодирования служит представление о нейроне-детекторе. Нейрон-детектор — высокоспециализированная нервная клетка, способная избирательно реагировать на тот или иной признак сенсорного сигнала. Такие клетки выделяют в сложном раздражителе его отдельные признаки. Разделение сложного сенсорного сигнала на признаки для их раздельного анализа является необходимым этапом операции опознания образов в сенсорных системах. Нейроны-детекторы были обнаружены в 60-е гг. сначала в сетчатке лягушки, затем в зрительной коре кошки, а впоследствии и в зрительной системе человека.
Информация об отдельных параметрах стимула кодируется нейроном-детектором в виде частоты потенциалов действия, при этом нейроны-детекторы обладают избирательной чувствительностью по отношению к отдельным сенсорным параметрам.
Виды нейронов-детекторов. Наиболее детально нейроны-детекторы исследованы в зрительной системе. Речь идет, в первую очередь, об ориентационно- и дирекционально-чувствительных клетках. За открытие феномена ориентационной избирательности нейронов зрительной коры кошки ее авторы Д. Хьюбел и Т. Визел в 1981 г. были удостоены Нобелевской премии. Явление ориентационной избирательности заключается в том, что клетка дает максимальный по частоте и числу импульсов разряд при определенном угле поворота световой или темновой полоски или решетки. В то же время при других ориентациях стимулов те же клетки отвечают плохо или не отвечают совсем. Эта особенность дает основание говорить об остроте настройки нейрона-детектора и предпочитаемом диапазоне реагирования. Дирекционально-избирательные нейроны реагируют на движение стимула, демонстрируя предпочтение в выборе направления и скорости движения (см. Хрестомат. 5.2).

Длинная и узкая полоска света вызывает реакцию сложной клетки независимо то того,
в каком месте рецептивного поля она предъявлена, если только ее ориентация оптимальна (три верхних записи).
Если ориентация полосы отличается от оптимальной, клетка реагирует слабее или не отвечает совсем (нижняя запись) (по Хьюбелу, 1991)
По своим способностям реагировать на описанные характеристики зрительных стимулов (ориентацию, скорость и направление движения) нейроны-детекторы делятся на три типа: простые, сложные и сверхсложные. Нейроны разного типа расположены в разных слоях коры и различаются по степени сложности и месту в цепи последовательной обработки сигнала.
Помимо этого, описаны нейроны-детекторы, которые реагируют в основном на стимулы, похожие на те, что встречаются в жизни, например, движущуюся тень от руки, циклические движения, напоминающие взмахи крыльев и т.д. Сюда же относятся нейроны, которые реагируют лишь на приближение и удаление объектов. Выделены также нейроны — детекторы цвета. Наиболее часто встречаются нейроны, чувствительные к синему цвету (с длиной волны 480 нм), зеленому цвету (с длиной волны 500 нм) и красному (с длиной волны 620 нм).
В высших центрах мозга обнаружены также зрительные нейроны, особо чувствительные к стимулам, сходным с человеческим лицом или какими-то его частями. Ответы этих нейронов регистрируются при любом расположении, размере, цвете "лицевого раздражителя". Важно отметить, что эти нейроны находятся не только в неокортексе, но и в более глубоких структурах мозга — в базальных ганглиях, таламусе и др. Иными словами, среди внешних стимулов есть наиболее "предпочтительные", такие, которые оказываются наиболее "удобными" для обработки нейронными механизмами восприятия.
Предполагается также, что существуют нейроны с возрастающей способностью к обобщению отдельных признаков объектов и полимодальные, т.е. обладающие способностью реагировать на стимулы разных сенсорныхмодальностей (зрительно-слуховые, зрительно-сомато-сенсорные и т.д.).
Описаны нейроны-детекторы и в других сенсорных системах: слуховой и соматосенсорной. В первом случае речь идет о детектировании положения источника звука в пространстве и направления его движения. Во втором, активность нейронов детекторов связана с определением движения тактильного стимула по коже или величиной суставного угла при изменении положения конечности.
Несмотря на то, что имеющихся данных о механизмах детектирования и в зрительной, и особенно в других модальностях (слуховой, соматосенсорной, обонятельной) явно недостаточно, тем не менее многие исследователи в настоящее время рассматривают принцип нейронного детектирования как универсальный принцип строения и функционирования всех сенсорных систем.
Обобщенная модель сенсорной системы. Детекторный принцип кодирования положен в основу "обобщенной модели сенсорной системы, выполняющей активный синтез при внутреннем отображении внешнего стимула" (Соколов, Вайткявичус, 1989). Модель воспроизводит все этапы процесса переработки информации от возникновения возбуждения на выходах рецепторов до формирования целостного образа.
Преобразование информационного потока в ней осуществляется с помощью нескольких типов формальных нейронов (детекторов, гностических нейронов, нейронов-модуляторов, командных, мнемических и семантических нейронов), связанных между собой стабильными и пластическими связями двух типов: информационными и модулирующими.
Предполагается, что внешний раздражитель через органы чувств создает распределенное возбуждение на выходе рецептора. В результате первичного анализа из этого потока возбуждения выделяются отдельные признаки стимула. На следующем этапе происходит организация целостного образа, в ходе этой стадии в зрительной системе человека по отдельным фрагментам возникает гипотеза о том, что это может быть. Гипотетические представления об объекте (ожидаемый образ) извлекается из памяти и сопоставляется с той информацией, которая поступает из сенсорной системы. Далее принимается решение о соответствии или несоответствии гипотезы объекту, проверяются уточняющие гипотезу признаки.
Концепция частотной фильтрации. Применительно к работе зрительной системы описан и другой предполагаемый механизм восприятия: частотная фильтрация. Предполагается, что зрительная система, в первую очередь кора мозга, настраивается на восприятие пространственной информации разного частотного диапазона. Другими словами, допускается, что зрительная система человека содержит нейронные комплексы, которые наделены свойствами двумерных пространственно-частотных фильтров, осуществляющих анализ параметров стимула по принципу, который описывается разложением Фурье (см. тему 2 п. 2.1.1). Причем существует, по-видимому, множество относительно "узких" фильтров, настроенных на восприятие разных пространственных частот.
Преимущество системы, основанной на частотном анализе, состоит в том, что она упрощает узнавание знакомых объектов, которые имеют увеличенные или уменьшенные размеры. При этом предполагается, что в системе памяти фиксируется только гармонический состав (перечень волновых составляющих, полученный в результате разложения), он не зависит от реального размера объекта и это делает опознание стимула более экономным.
Дискуссионные вопросы. Обе концепции (детекторная и частотной фильтрации) не дают четкого ответа на один вопрос: как из некоторого числа элементов, будь это реакции нейронов-детекторов или коэффициенты Фурье, создается целостный образ, т.е. неопределенным остается механизм визуального синтеза и константности зрительного образа.
Один из основателей детекторной концепции Д. Хьюбел так характеризует эту проблему: "…часто спрашивают, каковы наиболее правдоподобные предположения относительно того, каким образом распознаются видимые объекты, становятся ли клетки при переходе к более центральным уровням все более специализированными, так что на каком-то уровне могут найтись клетки, реагирующие на лицо одного-единственного конкретного человека — например, чьей-то бабушки? Такое представление, называемое "теорией бабушкиной клетки", вряд ли можно принимать всерьез. Можем ли мы обнаружить отдельные клетки для бабушки улыбающейся, плачущей или занимающейся шитьем? Или отдельные клетки, отражающие понятие или определение "бабушки"? И если бы у нас действительно имелись "бабушкины клетки", куда они посылали бы свои выходные сигналы? (Д. Хьюбел, 1991. С. 228).
Нейронный ансамбль как основа перцепции. В качестве возможного решения предлагается следующий механизм. Воспринимаемый объект активирует определенную группу клеток — "нейронный ансамбль", каждый член которого может принадлежать также к другим ансамблям. Поскольку известно, что разрушение небольшого участка мозга обычно не ведет к исчезновению определенных воспоминаний, приходится предполагать, что клетки одного ансамбля не сосредоточены в одной корковой зоне, а разбросаны по многим зонам. Таким образом, "бабушке", занимающейся шитьем", должен соответствовать более крупный ансамбль, включающий бабушку по определению, бабушкино лицо и процесс шитья. Предлагаемая схема, однако, носит скорее гипотетический характер.
Наряду с этим, экспериментально доказано существование специализированных нейронов в мозге человека. Многочисленные данные такого рода были получены в клинических исследованиях Н.П. Бехтеревой с сотрудниками (1985, 1988). Более того, ими был сформулирован общий психофизиологический принцип, в соответствии с которым кодирование содержания психической деятельности осуществляется комбинациями частот импульсной активности в паттернах разрядов нейронов мозга и в характеристиках их взаимодействия. В частности, например, было показано, что паттерны текущей частоты разрядов нейронов некоторых структур мозга при восприятии вербальных стимулов способны отражать акустические и общие смысловые характеристики слов.
Тем не менее в исследованиях восприятия, выполняемых с помощью регистрации активности нейронов, проблема формирования и опознания образа остается мало изученной (см. Видео).
Изучение физиологических основ восприятия проводится с помощью макропотенциалов мозга: фоновой электроэнцефалограммы, а также вызванных и событийно-связанных потенциалов, причем использование ЭЭГ имеет два разных аспекта. Первый связан с выявлением параметров ЭЭГ, имеющих отношение к кодированию стимула. Второй, более традиционный, представляет собой изучение параметров ЭЭГ отдельных зон коры или показателей их совместного взаимодействия при восприятии стимулов.
ЭЭГ — показатели восприятия. Среди ритмов ЭЭГ наибольшее внимание исследователей в этом плане привлекает альфа-ритм (8-12 кол/с), который регистрируется преимущественно в задних отделах коры в состоянии спокойного бодрствования. Известно, что при предъявлении стимулов имеет место подавление или "блокада" альфа-ритма: причем она тянется тем больше, чем сложнее изображение.
Существует немало предположений, касающихся роли альфа-ритма в обеспечении активности мозга как целого. Основоположник кибернетики Н. Винер и вслед за ним ряд других исследователей считали, что этот ритм отражает механизм временного сканирования ("считывания") информации и тесно связан с механизмами восприятия и памяти. Действительно, по некоторым данным, биопотенциальное поле альфа-ритма связано с функцией сканирования информации в реальном времени и тем самым связано с процессами межмодальной сенсорной интеграции в процессах восприятия и памяти. По другим представлениям, альфа-ритм связан с действием селектирующих механизмов мозга, выполняющих функцию резонансного фильтра и, таким образом, регулирующих поток сенсорных импульсов.
Поиск электрофизиологических показателей восприятия в параметрах альфа-ритма опирается на представление о том, что воспринимаемая человеком информация кодируется комбинациями фаз и частот периодических нейронных процессов, которые находят свое отражение в характеристиках альфа-ритма. Об этом говорит сайт https://intellect.icu . Предполагается, что такая особенность ЭЭГ, как пакет волн создается синхронизированной когерентной активностью группы нейронов, расположенных в разных участках мозга и образующих ансамбль. Предположительно все волны одного пакета хранят информацию об одном образе или его части и при восприятии опознается только тот образ, который закодирован ритмической активностью наибольшего числа нейронов в каждый данный момент времени (А.Н. Лебедев, 1985).
Экспериментально было показано, что значения параметров колебаний системы нейронов, оцениваемые, в частности, по расстоянию между соседними спектральными пиками внутри альфа-диапазона, могут служить аргументами уравнений, предсказывающих некоторые особенности восприятия. Например, чем больше период доминирующих колебаний в ЭЭГ человека и чем больше разнообразие воспринимаемых и ожидаемых стимулов, тем медленнее осуществляется их восприятие.
Наряду с этим существуют исследования пространственно-временных отношений потенциалов мозга при восприятии сенсорной информации. В отличие от предыдущего этот подход учитывает два фактора: время восприятия и его мозговую организацию. Речь идет о множественной регистрации ЭЭГ из разных зон коры больших полушарий в процессе восприятия. Поскольку нейронные ансамбли, участвующие в переработке информации распределены по разным отделам мозга, в первую очередь коры больших полушарий, логично считать, что перцептивный акт будет сопровождаться изменением пространственного соотношения ЭЭГ. Действительно изучение дистантной синхронизации биопотенциалов коры (М.Н. Ливанов, 1977) в ходе зрительного восприятия позволяет выявить следующее: вначале наблюдается преимущественная активация задних отделов коры обоих полушарий, затем в процесс вовлекаются передние отделы правого полушария. Узнавание связано с активным включением в процесс центральных и фронтальных зон коры.
Реорганизация биоэлектрической активности в процессе перцептивного акта характерна для всех видов чувствительности. Однако в зависимости от вида анализатора она имеет разную пространственную картину. Как правило, на ранних этапах процесса наибольшую активность демонстрирует проекционная зона (зрительная, слуховая, соматосенсорная), на завершающих этапах в процесс включаются передние отделы коры.
Вызванный потенциал как коррелят перцептивного акта. Наиболее адекватным инструментом изучения физиологической природы перцептивного процесса является метод регистрации вызванных (ВП), или событийно-связанных потенциалов. Последнее обусловлено тем, что, во-первых, ВП — это реакция мозга на внешнее раздражение, по длительности сопоставимая со временем перцептивного акта, во-вторых, ВП, взятые отдельно в каждой зоне коры, имеют свои характеристики, обусловленные функциональными особенностями этой зоны. Таким образом, при предъявлении стимула любой модальности (зрительного, звукового, тактильного и др.) в разных зонах коры возникают специализированные реакции, сопоставимые по длительности с реальным психологическим процессом восприятия.
Первые исследования в этом направлении ставили своей целью соотнести конфигурацию и параметры ВП с типом стимула и выделить варианты ответов, связанных с содержанием и физическими характеристиками стимула, например, установить отвечает ли мозг специфическим ответом на геометрические фигуры (круг, квадрат) или определенные слова. Предпринимались даже попытки определять по конфигурации ВП, какой стимул предъявлялся испытуемому. Несмотря на определенные положительные результаты, эти исследования не получили развития.
На первый план выступил иной подход. В его основе лежали представления о том, что компонентная структура ВП отражает не конкретное содержание или признак воспринимаемого стимула, а некоторые общие последовательные операции головного мозга по обработке стимула.
ВП и последовательная модель переработки информации. Было установлено, что в конфигурации ВП можно выделять компоненты двух типов: ранние специфические (экзогенные) и поздние неспецифические (эндогенные) компоненты. Обработка физических параметров стимула (сенсорный анализ) связывается с деятельностью специфических сенсорных систем и находит свое отражение преимущественно в параметрах экзогенных компонентов ВП. В зрительной модальности эти компоненты регистрируются в интервале до 100 мс от начала ответа, в слуховой и соматосенсорной модальностях — еще раньше, в пределах 10-50 мс от начала стимуляции. Эндогенные компоненты ВП, возникающие позднее и длящиеся до окончания ответа, предположительно отражают этапы более сложной обработки стимула: формирование образа, сличение его с эталонами памяти, принятие перцептивного решения.

Выделение когнитивной составляющей ответа:
а - ответ в условиях опознания значимого стимула у здорового испытуемого (45 лет): P1-N1-P2 собственно ответ на стимул, N2 - 170 мс и P3 - 357 мс;
б. Ответ на тот же стимул без опознания;
а-б. Разность ответов в условиях распознавания и в стандартных условиях без распознавания. Когнитивный комплекс.
Восприятие начинается на 76 мс, опознание и дифференцировка - компонент N2 - 170 мс,
принятие решения и запоминание - компонент Р3 - 354 мс. (по В.В.Гнездицкому, 1997)
Иллюстрацией такого подхода может служить концепция "информационного синтеза" А.М. Иваницкого (1986), согласно которой ранние компоненты ВП в интервале 0-100 мс связываются с оценкой физических параметров стимула, в среднелатентных компонентах в интервале 100-200 мс находят отражение процессы синтеза текущей сенсорной информации с содержанием памяти, а в поздних компонентах ВП отражается когнитивная оценка стимула как такового.

Вызванные потенциалы (ВП) мозга, возникающие в ответ на световой или кожный раздражитель (по А.М.Иваницкому, 1999).
Видно, что ранние волны соотносятся с чувствительностью, поздние - с мотивацией, значимостью стимула, а промежуточные - одновременно с двумя этими психофизическими показателями. Стрелкой на временной шкале обозначает момент предъявления стимула
Эти представления хорошо согласуются с нейрокибернетическими построениями Д. Эделмена и В. Маунткасла (1981), согласно которым повторный возврат возбуждений является мозговой основой всех психических процессов. Так, например, момент возникновения образа совпадает по времени со встречей в коре двух информационных потоков, один из которых представляет собой внешний сигнал, а другой память. Предполагается, что эта память и есть возникающая в сознании человека частица "я", по отношению к которой сигнал воспринимается как нечто внешнее (см. Хрестомат. 5.3).
Изложенные выше представления объяснимы с помощью последовательной модели переработки информации, согласно которой каждая стадия перцептивного процесса наступает не ранее, чем реализуется предыдущая.
В то же время имеются данные, трудно объяснимые в контексте последовательной модели переработки информации. Например, при регистрации зрительных ВП на особо сложные стимулы параметры этих стимулов могут обрабатываться во всем временном диапазоне ВП. Последнее предполагает наличие иных моделей переработки информации: параллельной и циклической. Причем в рамках обоих моделей ВП имеют различное толкование.
В основе параллельной модели лежит представление о существовании в зрительной системе нескольких параллельно работающих подсистем, в которых переработка информации о разных признаках стимула происходит независимо, например, по одному каналу осуществляется описание формы объекта, по другому — контраста, цвета и т.д. В этой логике компоненты ВП рассматриваются как показатели включения разных каналов.
Таким образом, главное отличие параллельной модели от последовательной заключается в том, что отдельные компоненты здесь рассматриваются не как маркеры реализации конкретных стадий обработки информации, а как маркеры включения разных каналов обработки информации. Так, например, компонентная структура ВП на шахматные поля и решетки интерпретируется как последовательность включения разных каналов, обрабатывающих информацию о яркости (ярко-синий канал) и о пространственном контрасте, анализ которого необходим для выделения формы (канал формы).
При этом различия в выраженности относительно ранних и поздних компонентов ВП в ответах на низкие и высокие пространственные частоты отражают не временную последовательность процессов обработки высоких и низких пространственных частот, а различную пространственную локализацию генераторов ВП на разные пространственные частоты (Зислина, 1978). В принципе эти предположения имеют определенные основания, поскольку опираются на электрофизиологические данные, касающиеся источников генерации ВП, а те подтверждают, что относительно ранние компоненты ВП на шахматные поля и решетки с латентным периодом 75-125 мс генерируются в стриарной коре, а более поздние имеют своим источником парастриарную область.
Циклическая модель предполагает, что в перцептивном процессе могут существовать отдельные и независимые перцептивные циклы с разной внутренней структурой и продолжительностью отдельных стадий. При циклической обработке информации, по-видимому, происходит повторное активирование одних и тех же нейронных популяций. Переработка информации при этом обеспечивается не только однократными специфическими реакциями нейронов, но и их повторным вовлечением в фазический процесс.
При трудной перцептивной задаче правильное опознание с большей вероятностью происходит на поздних этапах переработки информации. По мере привыкания возрастает вероятность опознания сигнала на более ранних фазах переработки. При этом предполагается, что число компонентов ВП определяется количеством фаз переработки информации в корково-подкорковых нейронных сетях (Переслени и др., 1987). Таким образом, в одном и том же интервале времени после предъявления одних и тех же стимулов могут возникать различающиеся по числу компонентов и конфигурации ВП. И наоборот, внешне одни и те же компоненты ВП могут соответствовать разным стадиям обработки стимула.
Эти представления ставят под сомнение возможность существования фиксированной схемы обработки информации по типу "постоянный стимул — неизменная реакция". В то же время логически непротиворечивое обоснование описанные выше факты получают с позиций гипотезы когнитивных детерминант организации перцептивного акта. Согласно этой гипотезе, порядок следования и набор операций (и соответствующих компонентов ВП) в каждом перцептивном акте непостоянен (от пробы к пробе) и определяется в процессе поступления информации и по ходу ее обработки. Решающую роль при этом будут играть условия задачи и опыт субъекта в организации информации.
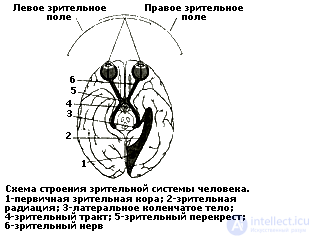
В основе этого подхода лежит представление о системном характере взаимодействия структур мозга в обеспечении психических функций (Л.С. Выготский, А.Р. Лурия, Е.Д. Хомская, М.Н. Ливанов, О.С. Адрианов и др.). Проблема участия разных отделов мозга, в первую очередь коры, в обеспечении восприятия изучается экспериментально с помощью разных методов: электроэнцефалографии и вызванных потенциалов, компьютерной томографии, прямого раздражения коры мозга, анализа нарушений восприятия при очаговых поражениях мозга.
Функциональное обеспечение восприятия связано с деятельностью второго блока. Морфологически он представлен задними отделами коры больших полушарий и включает "аппараты" зрительной (затылочной), слуховой (височной), общечувствительной (теменной), а также соматосенсорной (постцентральной) зон коры. Все перечисленные "аппараты" построены по общему принципу: они включают первичные проекционные зоны ("корковые концы анализаторов") и ассоциативные вторичные и третичные зоны.
Общей особенностью первичных зон является топическая организация, т.е. каждому участку такой зоны соответствует определенный участок периферической рецепторной поверхности. Проекция сетчатки образует ретинотопическую организацию затылочной зоны, проекция кортиева органа — тонотопическую слуховой зоны, проекция рецепторной поверхности кожи — соматотопическую соматосенсорной зоны. Такой принцип организации обеспечивает своеобразное картирование сетчатки, кортиева органа и кожной поверхности.
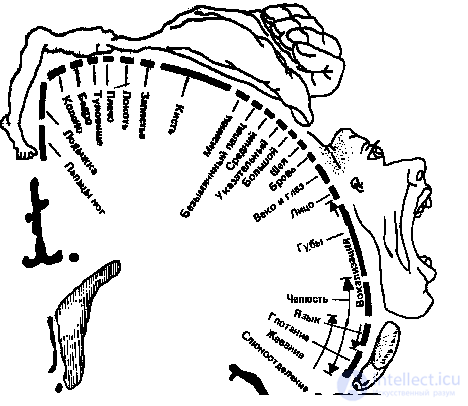
Лучше других изучена соматотопическая организация соматосенсорной зоны. Хорошо известен соматосенсорный гомункулус - схематический "человечек", отражающий пространственную представленность разных участков тела в этой зоне коры. Фактически он представляет карту этой зоны, которая была получена во время операций на мозге человека с помощью прямой электрической стимуляции. Такая стимуляция вызывает у человека ощущения, которые распределены по разным участкам тела, но неравномерно — более всего представлены в коре функционально наиболее значимые участки рецепторной поверхности (лицо, язык, рука).
Последнее справедливо для всех проекционных зон, так, например, в затылочной зоне большую часть занимает проекция центрального участка, где преимущественно сосредоточены колбочки, обеспечивающие высокую контрастную чувствительность и цветное зрение.
Таким образом, при восприятии стимула не происходит равномерно возбуждения всей проекционной зоны: активируются, главным образом, те нейроны, к которым поступает информация от возбужденных периферических элементов — рецепторов. Этот первичный этап нередко обозначают как сенсорный анализ.
Принципиально меняется характер обработки информации при переходе к вторичным ассоциативным зонам коры. Являясь модально специфическими, эти зоны, по образному утверждению А.Р. Лурии, превращают топическую проекцию раздражения в функциональную организацию. Это значит, что обработка информации в ассоциативных зонах происходит на другой основе. Опираясь на представления, сформулированные в разделе п. 5.1, можно предположить, что при переходе информации от первичной проекционной зоны к ассоциативным изменяется природа кодов, в которых обрабатывается информация. За счет каких именно механизмов осуществляется реорганизация потоков импульсной активности нейронов пока неизвестно. Выше (см. п. 5.2) говорилось о существовании нейронов разной специализации (мнемических, семантических, гностических и др.), нейронных ансамблей, осуществляющих функции обобщения при восприятии. Однако недостаточно экспериментальных данных для описания нейронных процессов, сопровождающих обработку информации во вторичных зонах коры. Тем не менее деятельность этих зон связывается с интеграцией разобщенных возбуждений первичной проекционной коры и формированием целостных образов восприятия. Функции третичных зон связаны с процессами межмодального обобщения и построения картинны мира.
Процесс обработки информации имеет много этапов и начинается на уровне рецепторов: чувствительных элементов кожи, сетчатки, кортиева органа в улитке и т.д. Кроме того, каждый анализатор включает целый ряд подкорковых переключений - звеньев (в частности, например, на уровне таламуса), где проводится начальная обработка информации. Только после этих стадий (т.е. осуществляемых на подкорковом уровне) уже частично обработанная информация поступает по проводящим путям в проекционные зоны коры и из них уже во вторичные и третичные. Такая общая последовательность стадий имеет свою специфику для каждого вида чувствительности.
Так, в зрительном анализаторе выделяются две системы обработки информации о внешнем мире. Первая из них ответственна за опознание объекта, вторая определяет локализацию объекта во внешнем зрительном поле. Выполняя разные функции зрительного восприятия, эти системы отличаются по своему морфологическому строению. Хотя обе системы начинаются в сетчатке, но первая система берет начало от так называемых клеток типа Х, вторая система — от клеток типа У. Далее эти системы имеют свое представительство в разных подкорковых центрах: первая система продолжает обработку информации о форме объекта в латеральных коленчатых телах таламуса, откуда информация поступает в зрительные центры коры: первичную проекционную зону, вторичные ассоциативные поля, и оттуда в нижневисочную зону коры. При выборочном поражении отдельных участков перечисленных зон страдает восприятие отдельных качеств объекта: только цвета, только формы или движения. Интеграция всех раздельно обрабатываемых признаков объекта происходит в нижневисочной коре, которая отвечает за окончательное формирование целостного зрительного образа объекта.
Вторая система имеет иную мозговую топографию. От сетчатки волокна этой системы направляются в собственные подкорковые центры — верхние двухолмия. В этой структуре осуществляется не только зрительное восприятие пространственных характеристик объекта, в ней же находятся центры, управляющие движением глаз. Активация этих центров запускает саккаду — быстрое скачкообразное движение глаз, амплитуда и направление которой обеспечивает попадание стимула в центральное поле зрения. Это необходимо, потому что именно центральное поле зрения обладает наилучшими способностями к восприятию высококонтрастных сложных стимулов. Дальнейшая обработка информации в этой системе связана с деятельностью другого таламического ядра - так называемой подушки и теменной области коры больших полушарий. В этой области происходит интеграция информации от первичной зрительной коры и центров, контролирующих движения глаз. Слияние двух потоков информации создает константный, т.е. постоянный экран внешнего зрительного поля. Благодаря этому перемещающийся по сетчатке во время движения глаз образ зрительного мира остается неизменным.
Таким образом, на основе взаимодействия информации, поступающей из разных отделов зрительной системы и системы управления глазодвигательной активностью, осуществляется построение целостной и стабильной картины миры.
Модель зрительного восприятия, учитывающую морфологическое строение анализатора, предложил В.Д. Глезер (1985). Он выделяет четыре уровня переработки зрительной информации: от зрительных центров подкорки, реализующих элементарный сенсорный анализ, — через проекционную кору, где осуществляется пространственно-частотный анализ, к третьему уровню, на котором происходит сегментация зрительного пространства на отдельные фигурные элементы, и от него — к высшим зрительным центрам нижневисочной и заднетеменной областям коры, осуществляющим опознание и обучение, — таков путь преобразования и описания зрительной информации по схеме Глезера.
Предложенная модель далеко не единственная в описании предполагаемых механизмов, обеспечивающих целостность зрительного восприятия. Проблема состоит в том, что каждый анализатор имеет вертикально организованные звенья (расположенные на пути от рецептора к первичной проекционной зоне), горизонтально организованные звенья (первичные, вторичные и третичные зоны коры) и латерально организованные звенья (симметричные подкорковые образования и зоны коры в левом и правом полушарии). Однако в подавляющем большинстве случаев предлагаемые модели ограничиваются одной или двумя из перечисленных составляющих.
Принято подразделять цитоархитектонические поля коры на первичные, вторичные и третичные (рис. 1).

Рис. 1 Локализация первичных и вторичных корковых полей. Первичные чувствительные поля заштрихованы, первичное двигательное поле черное. Вторичные чувствительные поля выделены мелким, вторичные двигательные поля---крупным пунктиром. Третичные поля коры оставлены белыми. Нумерация корковых полей по Бродману.
Первичные поля коры представляют собой четко отграниченные участки, которые соответствуют центральным частям анализаторов. В эти поля проходит по специфическим проекционным афферентным путям основная масса сигналов от органов чувств. Первичные поля характеризуются сильным развитием внутренней зернистой пластинки. Первичные поля связаны с реле-ядрами таламуса и ядрами коленчатых тел. Они имеют экранную структуру и, как правило, жесткую соматотопическую проекцию, при которой отдельные участки периферии проецируются в соответствующие им участки коры. Повреждение первичных полей коры сопровождается нарушением непосредственного восприятия и тонкой дифференцировки раздражений.
Представительство в коре внутренних органов изучено недостаточно, по-видимому, оно имеет более диффузный характер. Важная роль в регуляции функций внутренностей отводится лимбической области коры, в которую входит поясная и парагиппокампальная извилины, гиппокамп, прозрачная перегородка и подмозолистое поле. Лимбическая кора вместе с подкорковыми образованиями (миндалевидное тело, ядро поводков, ядра сосцевидных тел) составляет лимбическую систему, которая представляет субстрат эмоций и реакций, связанных с основными биологическими влечениями (голод, жажда, страх и т.д.).
Вторичные поля коры примыкают к первичным полям. Их можно рассматривать как периферические части корковых анализаторов. Эти поля связаны с ассоциативными ядрами таламуса. При поражении вторичных полей сохраняются элементарные ощущения, но нарушается способность к более сложным восприятиям. Вторичные поля не имеют четких границ, в них не выражена соматотопическая проекция.
Вторичное поле общей чувствительности локализуется в верхней теменной дольке (поля 5, 7). Вторичные зрительные поля (18, 19) занимают медиальную поверхность затылочной доли и большую часть латеральной поверхности. Вторичное слуховое поле (22) находится в верхней и средней височных извилинах. Вторичные обонятельные и вкусовые поля локализуются в парагиппокампальной извилине и крючке (поля 28, 34).
Третичные поля коры отличаются наиболее тонкой нейронной структурой и преобладанием ассоциативных элементов. Они занимают всю нижнюю теменную дольку и часть верхней теменной дольки, а также затылочно-височно-теменную область. Эти поля связаны с задними ядрами таламуса. В третичных полях осуществляются наиболее сложные взаимодействия анализаторов, лежащие в основе познавательного процесса (гнозия), формируются программы целенаправленных действий (праксия).
Кора височной доли имеет отношение к хранению и воспроизведению впечатлений. При электрическом раздражении некоторых точек височной коры наблюдаются своеобразные реакции в форме «вспышек пережитого» или ощущения «уже виденного». Полагают, что в коре височных долей создается нейронная запись потока сознания, она хранится неопределенно долгое время, но не может произвольно воспроизводиться, а «оживает» лишь при искусственной стимуляции и некоторых болезненных состояниях.
Особенности функциональной специализации левого и правого полушарий мозга широко исследуются. Подробный анализ этой проблемы представлен в книгах Н.Н. Брагиной и Т.А. Доброхотовой (1981, 1994), Е.Д. Хомской (1987), С. Спрингер и Г. Дейча (1983) и многих других.
Нервная система человека устроена таким образом, что каждое полушарие мозга получает информацию главным образом от противоположной стороны тела. Этот принципконтралатеральной проекции относится как к общей телесной, тактильной чувствительности, так и к зрению и слуху, хотя применительно к последним картина не столь однозначна. Однако использование соответствующих методик позволяет избирательно подавать информацию только в одно полушарие и выявлять тем самым значительные различия в функциональных способностях двух полушарий.
Наиболее изучены межполушарные отношения при зрительном и слуховом восприятии. Особенности специализации полушарий при восприятии звуковых стимулов, в первую очередь речи, представлены в теме 8 (см. п. 8.4; п. 8.5), поэтому здесь основное внимание будет посвящено специализации полушарий мозга при зрительном восприятии.
Как показывают клинические исследования, разрушение центральных зрительных зон в одном из полушарий ведет к утрате противоположной половины поля зрения (правой при левостороннем повреждении и левой — при правостороннем). Зная место повреждения зрительной коры, можно предсказать, какова будет потеря зрения. Однако предсказать реакции человека на такое повреждение сложно, поскольку нервная система человека обладает компенсаторными возможностями.
Среди гипотез относительно природы межполушарных различий при зрительном восприятии распространены две: одна связывает эти различия с вербализацией воспринимаемых стимулов, по другой — различия коренятся в особенностях стиля работы каждого полушария — аналитического для левого и целостного глобального для правого. В целом доминирует точка зрения, что превосходство того или иного полушария при восприятии зрительных стимулов определяется соотношением двух этапов переработки: зрительно-пространственного, в котором преобладает правое полушарие, и процессов вербализации, реализуемых левым.
Предпринимаются попытки объяснить межполушарные различия на основе специализации нейронов в перцептивном процессе. Известной в этом плане является модель, приписывающая полушариям головного мозга определенную специализацию относительно обработки разных пространственных частот. По этим предположениям по-разному осуществляется частотная фильтрация в зрительных центрах того и другого полушария. Правое полушарие с большим успехом обрабатывает низкие пространственные частоты и, следовательно, в большей степени связано с восприятием контуров объектов и их крупных деталей. Левое, напротив, более успешно обрабатывает высокие частоты, и в большей степени ответственно за восприятие мелких деталей изображения.
В наиболее общем виде специализация полушарий в обеспечении зрительного восприятия приведена в таблице 5.2.
Таблица 5.2. Различия между полушариями при зрительном восприятии
| Левое полушарие | Правое полушарие |
|---|---|
| Лучше узнаются стимулы | |
| Вербальные Легко различимые Знакомые |
Невербальные Трудно различимые Незнакомые |
| Лучше воспринимаются задачи | |
| Оценка временных отношений Установление сходства Установление идентичности стимулов по названиям Переход к вербальному кодированию |
Оценка пространственных отношений Установление различий Установление физической идентичности стимулов Зрительно-пространственный анализ |
| Особенности процессов восприятия | |
| Аналитичность Последовательность (сукцессивность) Абстрактность, обобщенность, инвариантное узнавание |
Целостность (гештальт) Одновременность (симультанность) Конкретное узнавание |
| Предполагаемые морфофизиологические различия | |
| Фокусированное представительство элементарных функций | Диффузное представительство элементарных функций |
В целом следует заключить, что правое "пространственное" и левое "временное" полушария обладают своими специфическими способностями, позволяющими им вносить важный вклад в большинство видов когнитивной деятельности. По-видимому, у левого больше возможностей во временной и слуховой сферах, а у правого в пространственной и зрительной. Эти особенности, вероятно, помогают левому полушарию лучше отмечать и обособлять детали, которые могут быть четко охарактеризованы и расположены во временной последовательности. В свою очередь единовременность восприятия пространственных форм и признаков правым полушарием, возможно, способствует поиску интегративных отношений и схватыванию общих конфигураций. Если такая интерпретация верна, то, по-видимому, каждое полушарие перерабатывает одни и те же сигналы по-своему и преобразует сенсорные стимулы в соответствии со специфичной для себя стратегией их представления.
Восприятие мира человеком осуществляется посредством его сенсорных систем при этом обрабатывается поток информации, скорость которого составляет около 11 миллионов бит в секунду.
Восприятия возникают в результате обработки сенсорной информации из которой извлекается (формируется) семантическая, прагматическая и другая информация. В случае, если сформированные воспринятия (семантические содержания) недостаточны сильны для того, чтобы достичь сознание (так называемые подпороговые (сублиминальные) восприятия), они могут сохраниться в личном бессознательном , из которого в дальнейшем могут быть извлечены в область сознания, например, с применением гипноза.
Визуальное восприятие мира осуществляется посредством зрительной системы и хотя визуальная картина мира кажется цельной, она собирается из результатов нервной деятельности нескольких десятков взаимодействующих областей мозга, специализированных на реализацию конкретных аспектов зрения. К 2000 году было идентифицировано более 30 участков коры головного мозга, связанных с глазами через зрительную зону V1 и выполняющих специфические функции обработки визуальной информации. При формировании визуальной картины мира зрительная система человека обрабатывает поток информации в 10 миллионов бит в секунду.
Зрительная и пространственная информация выделяется из визуальной информации, находящейся в сенсорной иконической памяти (см. память), системами распознавания — «что» (по вентральному пути) и локализации — «где»(по дорсальному пути) выявляется семантическая информация: зрительная о свойствах объектов (о форме, цвете и распределении объектов) и пространственная (о местоположении и движении объектов).
С рождения младенцы интересуются человеческими лицами, однако имеют очень приблизительную модель лица и потому смотрят почти на любой круглый предмет, имеющий два «глаза» и «рот» и находящийся на расстоянии, приблизительно 20 см . К четырем или пяти месяцам дети начинают уверенно отличать лица от других предметов. Вероятно, что это связанно с развитием веретенообразной (веретеновидной) извилины, участка на границе между затылочной и височной долями, вентральная поверхность которой специализируется на распознавание лиц . По всей видимости, активизация веретенообразной извилины происходит уже у двухмесячных младенцев. При поражении этой области возникает прозопагнозия — расстройство восприятия лица, при котором теряется способность узнавать лица.
Обработка и восприятие визуальной информации о лице, осуществляется распределенной системой, состоящей из нескольких областей мозга . Ядро этой системы составляют: область в нижней затылочной извилине (OFA) , обеспечивающая начальный анализ отдельных частей лица; область в веретеновидной извилине (FFA), проводящая анализ инвариантных характеристик лица и обеспечивающая узнавание человека по лицу[10]; область в задней части верхней височной борозды (pSTS), активирующаяся при анализе изменчивых аспектов — выражения лица, движений губ при речи и направления взгляда[11]. В расширенной системе происходит дальнейший анализ направления взгляда (межтеменная борозда — IPS), семантики (нижняя лобная извилина — IFG, передняя височная кора — ATC), эмоциональной составляющей (миндалина — Amy, островковая кора — Ins), биографической (предклинье — PreCun, задняя поясная извилина — pCiG) и другой информации. Связанная с восприятием объектов, латеральная затылочная кора (LOC), может включаться в ранний анализ структуры изображения лица. При этом выявление различных аспектов визуальной информации о лице осуществляется не автономной работой отдельных областей мозга, реализующих конкретные функции, а взаимосвязанной скоординированной их работой[12].
Не вся информация, связанная с восприятием лиц может достигать сознания. Так, проведенные в 2004 и 2006 годах исследования, показали, что у пациентов, которым экспонировали испуганные лица представителей других рас, наблюдалось повышение активности миндалевидного тела, в то время, как информация об эмоциях лиц на показанных картинках уровня сознания не достигала
Усвоение и понимание письменной и устной речи осуществляется частью коры головного мозга, называемой областью Вернике.Одной из самых популярных моделей естественного чтения текста человеком на средней скорости по целым словам является E-Z Reader, разработанная на основе положений когнитивной психологии.
Соматосенсорной коры головного мозга является частью мозга , которая принимает и кодирует сенсорную информацию от рецепторов всего тела.
Аффективное прикосновение - это тип сенсорной информации, которая вызывает эмоциональную реакцию и обычно носит социальный характер. Такая информация на самом деле кодируется иначе, чем другая сенсорная информация. Хотя интенсивность аффективного прикосновения по-прежнему кодируется в первичной соматосенсорной коре, чувство приятности, связанное с аффективным прикосновением, активизируется больше в передней поясной коре головного мозга . Контрастное изображение, зависящее от повышенного уровня кислорода в крови (жирный шрифт), выявленное во время функциональной магнитно-резонансной томографии (фМРТ), показывает, что сигналы в передней поясной коре головного мозга, а также в префронтальной коре сильно коррелируют с оценками приятности аффективного прикосновения. Ингибирующийтранскраниальная магнитная стимуляция (ТМС) первичной соматосенсорной коры подавляет восприятие интенсивности аффективного прикосновения, но не приятности аффективного прикосновения. Таким образом, S1 не участвует напрямую в обработке социально-эмоциональной приятности прикосновения, но по-прежнему играет роль в распознавании местоположения и интенсивности прикосновения.
Хроноцепция относится к тому, как течение времени воспринимается и переживается. Хотя чувство времени не связано с конкретной сенсорной системой , работа психологов и нейробиологов указывает на то, что человеческий мозг действительно имеет систему, управляющую восприятием времени , состоящую из высоко распределенной системы, включающей кору головного мозга. , мозжечок и базальные ганглии . Один конкретный компонент мозга, супрахиазматическое ядро , отвечает за циркадный ритм.(обычно называемые «внутренними часами»), в то время как другие кластеры клеток, по-видимому, способны вести хронометраж в более коротком диапазоне, известном как ультрадианный ритм .
Один или несколько дофаминергических путей в центральной нервной системе, по- видимому, оказывают сильное модулирующее влияние на умственную хронометрию , особенно на интервальное время.
Восприятие людей и высших животных включает функцию непосредственного определения различных математических свойств и соотношений, в том числе количественных.
Люди и животные обладают непосредственным восприятием множественности, позволяющим им практически мгновенно сравнивать размеры различных групп объектов, так младенцы обладают способностью без расчетов определять соотношение размеров групп при количестве объектов в них 1:2. Взрослые могут определять более сложные соотношения 7:8. Другой универсальной способностью восприятия является субитизация, умение мгновенно определять количество объектов в небольших группах (до четырех). Детекторы, селективные к числу объектов в поле зрения, обнаружены в височной коре обезьян.
Исследования, проводимые с применением фМРТ показывают, что количественные значения активируют участки, расположенные в лобных и задних отделах теменных долей, головного мозга. Одним из ключевых мест является внутритеменная борозда, где представлен семантический смысл чисел. У людей, страдающих дискалькулией — неспособностью к изучению арифметики, данный участок мозга меньше, чем у здоровых людей, и недостаточно активен.
Есть предположение, что в мозге образ множества чисел представлен в виде прямой, точки которой в порядке возрастания соответствуют числам. Из-за этого время ответа, «какое из чисел больше», для близких чисел (например, 7 и 8), требует больше времени, чем для тех, разность которых велика (8 и 2).
Есть информация, что некоторые оленеводы могут мгновенно определять потерю нескольких голов в полутора, двухтысячных стадах оленей. При этом они не обладали умениями счета больших количеств.
Первые системные исследования развития непосредственного восприятия математических характеристик, в рамках изучения развития общих когнитивных способностей детей проводил французский психолог Пиаже.
Одним из давно известных парадоксов визуального восприятия является трудность определения направления движения объекта, занимающего большую часть зрительного поля, особенно, если скорость движения этого объекта велика. Недавно исследователи Московского государственного психолого-педагогического университета (МГППУ) выявили нейрофизиологические основы этого эффекта восприятия.
Ученые МГППУ исследовали нейронные механизмы, управляющие обработкой поступающей информации. Такие механизмы включают тормозные нейроны и функционируют на основе поддержания определенного баланса тормозящих и возбуждающих импульсов, управляющих восприятием информации. Часть таких механизмов обеспечивает сокращение объема обрабатываемой информации, что особенно критично для зрительной информации из-за высокой интенсивности ее поступления. Механизмы обеспечения сокращения объема обрабатываемой информации включают фильтрацию, отбора из огромного потока поступающей информации только тех ее составляющих, которые достаточны для формирования восприятия некоторой стандартизированной общей картины. Наличие таких механизмов необходимо, поскольку весь поток визуальной информации полностью обработать мозг не в состоянии.
Было выявлено, что функционирование таких механизмов проявляется в определенных свойствах гамма-волн, регистрируемых МЭГ и отражающих эффективность процессов нейронного торможения. При этом наибольшей информативностью обладают гамма-волны, вызываемые зрительными стимулами.
Функционирование нейронных механизмов обработки позволяет эффективно ориентироваться в среде для обычных условий, однако для некоторых нестандартных ситуациях, особенности их функционирования могут приводить к ошибкам. Так, например, чтобы четче определить направление движения в центре поля зрения, мозг тормозит обработку информации на периферии, сокращая ее восприятие. И в том случае, когда объект занимает большую часть зрительного поля, определить направление движения объекта, становится трудно, особенно, если скорость движения этого объекта велика.
1. Верно ли утверждение, что главное отличие параллельной модели работы зрительной системы от последовательной заключается в том, что в ней отдельные компоненты вызванных потенциалов рассматриваются как маркеры включения разных каналов обработки информации, а не как маркеры реализации конкретных стадий обработки информации?
Ответ 1,0,
2. Выберите правильный вариант ответа. Восприятие отдельных признаков объектов внешнего мира осуществляют:
Ответ 1,0,0,0
3. Выберите три правильных варианта ответа. При решении вопроса о природе того или иного нейронного кода выделяются такие главные вопросы:
Ответ 0,1,1,1,
4. Выберите правильный вариант ответа. Феномен ориентационной избирательности нейронов зрительной коры кошки открыли Д. Хьюбел и Т. Визел в:
Ответ 0,0,1,0,
5. Выберите правильный вариант ответа. Явление ориентационной избирательности заключается в том, что:
Ответ 0,0,1,
6. Выберите четыре правильных варианта ответа. В 'обобщенной модели сенсорной системы, выполняющей активный синтез при внутреннем отображении внешнего стимула' (Соколов, Вайткявичус, 1989) выделяется нескольких типов формальных нейронов:
Ответ 1,1,1,0,1,
7. Выберите правильный вариант ответа. Концепция частотной фильтрации в нейронных сетях основана на постулате двумерных пространственно-частотных фильтров, осуществляющих анализ параметров стимула по принципу, который описывается:
Ответ 0,0,0,1,
8. Выберите правильный вариант ответа. По представлениям Н. Винера, ритм ЭЭГ отражает механизм:
Ответ 0,1,0,0,
9. Выберите правильный вариант ответа. Компонентная структура вызванных потенциалов отражает:
Ответ 1,0,0,0,
10. Выберите правильный вариант ответа.Вызванные потенциалы рассматриваются как показатели:
Ответ 1,0,0,
11. Выберите правильный вариант ответа. Экзогенные компоненты в конфигурации вызванных потенциалов в зрительной модальности регистрируются в интервале:
Ответ 0,0,0,1,
12. Выберите три правильных варианта ответа. Экзогенные компоненты в конфигурации вызванных потенциалов отражают этапы сложной обработки стимула:
Ответ 1,1,1,0,
13. Выберите правильный вариант ответа. Согласно концепции 'информационного синтеза ' А.М. Иваницкого:
Ответ 0,0,1,0,
14. Выберите три правильных варианта ответа. В.Д. Глезер выделяет следующие уровни переработки зрительной информации:
Ответ 1,1,1,0,
К сожалению, в одной статье не просто дать все знания про психофизиология восприятия. Но я - старался. Если ты проявишь интерес к раскрытию подробностей,я обязательно напишу продолжение! Надеюсь, что теперь ты понял что такое психофизиология восприятия, сукцессивное восприятие, симультанное восприятие, потенциал действия, дирекционно-чувствительные клетки, эндогенный, экзогенный, симультанный, сукцессивный, поля коры и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Психофизиология
Ответы на вопросы для самопроверки пишите в комментариях, мы проверим, или же задавайте свой вопрос по данной теме.

Комментарии