Лекция
Привет, Вы узнаете о том , что такое корневое питание растений , Разберем основные их виды и особенности использования. Еще будет много подробных примеров и описаний. Для того чтобы лучше понимать что такое корневое питание растений , настоятельно рекомендую прочитать все из категории Агрохимия и биохимия.
Поглощение воды и питательных веществ через корни, т.е. корневое питание, тесно связано с углеродным питанием растений через листья. Хотя с энергетической точки зрения фотосинтез представляет собой процесс улавливания громадного количества солнечной энергии и превращения ее в потенциальную энергию урожая, успешно протекать он может только при оптимальных условиях корневого питания. Фотосинтез и корневое питание, в сущности, единый, хотя и многоступенчатый процесс поглощения растениями необходимых питательных веществ из внешней среды, превращения воды, углекислоты и Mинеральных солей в многочисленные органические соединения с утилизацией солнечной энергии и при участии ферментов. В состав растения входят почти все элементы периодической системы Д.И. Менделеева, присутствующие в почве и в воздухе. В них обнаружено более 75 химических элементов.
Минеральное (корневое) питание растений осуществляется через корни и включает в себя поглощение необходимых минеральных элементов из почвы. Эти минеральные элементы играют ключевую роль в различных биохимических процессах, росте и развитии растений. Важно, чтобы почва содержала достаточное количество и правильное соотношение этих элементов для обеспечения оптимального здоровья растений. Вот некоторые из основных минеральных элементов и их роли в корневом питании:
Азот (N): Азот является ключевым компонентом аминокислот, белков и других органических соединений. Он необходим для формирования зеленой массы, роста и развития растений. Недостаток азота может привести к замедленному росту и желтизне листьев.
Фосфор (P): Фосфор участвует в образовании нуклеиновых кислот, энергетических молекул (АТФ), а также в росте корней, цветении и формировании плодов.
Калий (K): Калий влияет на регулирование водного баланса, поддерживает структуру клеточных стенок, участвует в активации ферментов и регулирует фотосинтез.
Магний (Mg): Магний является составной частью хлорофилла, что делает его необходимым для фотосинтеза. Он также участвует в многих биохимических реакциях внутри растения.
Кальций (Ca): Кальций необходим для развития клеточных стенок, обеспечивает их прочность и устойчивость.
Сера (S): Сера входит в состав аминокислот, витаминов и многих других органических молекул. Он важен для белков и других биохимических процессов.
Микроэлементы (железо, цинк, медь, марганец, бор, молибден и др.): Эти микроэлементы нужны в небольших количествах, но играют критическую роль в различных физиологических процессах растений, таких как активация ферментов и участие в образовании биохимических соединений.
Управление минеральным питанием растений включает в себя поддержание оптимального уровня питательных веществ в почве, учет потребностей конкретных видов растений и регулярную мониторинг почвенных условий. Это позволяет обеспечить эффективное питание растений и поддержание их здоровья и роста.
В зависимости от содержания элементов в растениях их подразделяют на макроэлементы, микро- и ультрамикроэлементы. К макроэлементам относятся необходимые элементы, содержание которых в растениях варьирует от десятков до сотых долей процента (n% —1072%): C,H, О, №, P, К, Ca, Mg, №, $. К микроэлементам относятся: В, Cu, Zn, Mn, Мо, Co, Se и Fe. Их содержание в растениях колеблется в пределах 10°2- 10 -5%. Они входят в основном в состав многочисленных ферментов. Ультрамикроэлементы содержатся в растениях в количестве менее 107%. Их физиолого-биохимическая роль до настоящего времени четко не установлена. В малых количествах они присутствуют в воде, воздухе, солях используемых в качестве удобрений, в материале сосудов, используемых для проведения вегетационных опытов и других средах. К ультрамикроэлементам относятся Ag, Au, Cr, №, W, Br, U, Rb, Cs и др. Значение этих элементов в жизни растений очень мало изучено. Кроме перечисленных элементов растения поглощают и другие вещества, находящиеся в почве, которые, хотя и не являются необходимыми, но могут в одних случаях действовать на растения положительно, а в других — отрицательно. Об ультрамикроэлементах можно сказать, что никто экспериментально не доказал и никто не опроверг их физиологическую значимость для растений, поэтому к ним на таком же основании можно отнести все химические элементы
He вошедшие в группу макро- или микроэлементов. При сжигании органического вещества все элементы, кроме азота, остаются в золе, поэтому их часто называют зольными элементами. С водой в растение поступают кислород и водород. Незаменимыми являются такие элементы питания, без которых растения не в состоянии завершить свой жизненный цикл «от семени до семени». Эти элементы называются биогенные или биофильные. Каждый из них выполняет в растении определенную биохимическую и физиологическую роль. Отсутствие или острый недостаток необходимого элемента вызывает глубокие нарушения биохимических процессов обмена веществ, приводящих к морфологическим изменениям органов и гибели растениях. При выращивании растений в различных почвенно-климатических условиях потребность их в каждом питательном элементе неодинакова. Почти повсеместно для формирования высокого урожая сельскохозяйственных культур, прежде всего, требуются азот, фосфор и калий. Потребность в кальции возникает на кислых почвах со слабой буферностью и низкой степенью насыщенности основаниями. Высокое действие магния часто наблюдается на легких почвах дерново-подзолистой зоны. В настоящее время некоторые исследователи считают необходимым внесение в почву серы как элемента питания растений. Однако этот вопрос требует обстоятельного изучения, особенно в связи со значительным количеством серы, выпадающей на почву в промышленно-индустриальных центрах. Эффективность отдельных микроэлементов зависит от природных условий зоны.
4.1. Основные формы потребления элементов питания растениями (Кидин, 2008)
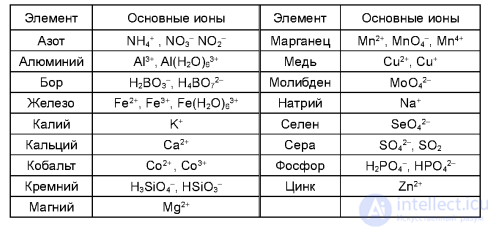
Положительное действие цинка, марганца и железа отмечается на нейтральных почвах степной зоны, особенно на карбонатных черноземах, а на почвах дерново-подзолистой зоны растения часто страдают от избытка этих элементов. В лесостепной и степной зонах редко можно наблюдать доказуемую прибавку урожая от применения меди как микроудобрения, за исключением некоторых опытов с кукурузой. На осушенных же болотных торфяных почвах нельзя получить хороший урожай зерновых культур без применения медных удобрений. Нельзя не отметить почти повсеместного положительного действия молибдена на урожай бобовых культур, что связано с участием этого элемента в физиолого-биохимических процессах фиксации молекулярного азота атмосферы клубеньковыми бактериями этих растений. Но эффективность этого элемента в различных почвенноклиматических условиях совершенно различна, что объясняется неодинаковым содержанием подвижных форм молибдена в почвах.
Необходим дифференцированный научный подход к разработке оптимальных условий питания растений и, конечно, специальное изучение роли каждого питательного элемента. В настоящее время состав и количество, а также формы соединений, в виде которых минеральные вещества должны даваться растениям, хорошо изучены (табл. 4.1). Об этом свидетельствует богатый опыт выращивания различных растений в водных и песчаных культурах и Ha гидропонике, где на питательных смесях из минеральных солей можно получить рекордно высокие урожаи. Растения могут для питания использовать и органические соединения: аминокислоты, органические кислоты, сахара, сахарофосфаты и др. При использовании азота аминокислот внутри растения происходит их дезаминирование, и освободившийся аммиак далее подвергается тем же превращениям, что и поглощенный корнями ион аммония. Значительно сложнее вопрос: как питаются растения? Несмотря на то, что изучению механизма поглощения ионов, их передвижению и превращению в растениях посвящено много исследований, в этом отношении еще много неясного (рис. 4.1). Большинство исследований по изучению роли питательных веществ и их превращения в растениях проводилось в условиях, когда складывался дефицит того или иного элемента путем исключения его из питательной среды.
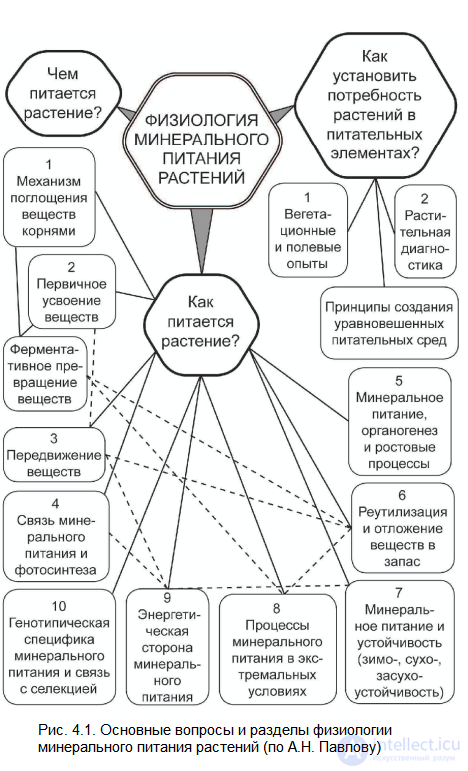
Рис. 4.1. Основные вопросы и разделы физиологии минерального питания растений (по А.Н. Павлову)
К концу ХХ столетия изучение процесса минерального питания: поглощения, передвижения веществ в растении — стали исследовать не только при дефиците того или иного элемента, но и при повышенном обеспечении растений элементами минерального питания. Это связано с необходимостью выяснения условий минерального питания, при которых реализуются потенциальные возможности продуктивности растения. Основные органы, с помощью которых растение питается, — это лист и корень. Последний выполняет несколько функций, прежде всего, поглощения минеральных веществ и воды из почвы. Другой важной функцией корня является переработка поступивших ионов: их восстановление и включение в различные органические соединения, в том числе и биосинтез физиологически активных соединений. У корней сильно развиты поверхность и длина периферийных, наиболее глубоко проникающих частей, но масса их незначительна. Благодаря им рассеянные в субстрате элементы минерального питания откладываются в верхних горизонтах почвы, где находится основная масса корней. Растение является, таким образом, не только потребителем почвенного плодородия, но и важнейшим его созидателем. От функции различных участков корня зависит целый ряд агротехнических приемов: глубина и характер заделки удобрений, глубина и ширина культивации при уходе за растениями, глубина основной обработки почвы и т.д. При прорастании зерновки трогается в рост главный зародышевый корень, затем появляются несколько новых зародышевых KOрешков. После начала кущения от стебля у основания листьев образуются узловые или придаточные корни, которые формируют вторичную корневую систему, выполняющую ту же функцию, что и зародышевые корни. Каждый корень имеет три основные зоны: 1) зону роста и растяжения длиной 1,5 мм — именно за счет деления клеток этой верхушечной меристемы происходит рост корня; 2) зону корневых волосков, или всасывания, характеризующуюся наличием особых выростов — корневых волосков длиной до 1 мм, а длина самой этой зоны 1-2 см; 3) зону боковых корней. В полевых условиях выращивания основное значение в питании растений принадлежит зоне корневых волосков, так называемой поглощающей зоне. Скорость поступления и передвижения питательных элементов в растении в сотни раз больше скорости таких физических явлений, как диффузия и осмос. Например, с помощью меченого углерода YC установлено, что углекислота из корней в листья передвигается за 10-15 мин. Скорость передвижения продуктов фотосинтеза из листьев в корни 40 - 100 см/ч. Еще быстрее поступают через корневую систему растений элементы питания, в том числе и внесенные в почву удобрения. Например, при погружении ячменя корнями в раствор, содержащий меченый фосфор 32P, его находили в листьях через 5 мин. Из корешков четырнадцатидневной кукурузы он поступал в листья через 2 мин. Такая же скорость передвижения питательных элементов из почвы в корни и листья растений наблюдалась в опытах с пшеницей и другими культурами. Об этом говорит сайт https://intellect.icu . Но вместе с тем скорость поглощения питательных веществ существенно изменяется с возрастом корня. Так, по мере старения растений кукурузы (с 20 до 80 дней) скорость поглощения №, P, К, Са и Mg уменьшается в десятки раз и более. Хотя молодые растения в абсолютных количествах поглощают во много раз меньше элементов минерального питания, чем взрослые, почва должна иметь высокое содержание этих элементов, чтобы достаточно удовлетворять высокую потребность корня в питательных веществах на ранних этапах его роста. При повышении концентрации питательных веществ в почве корни усиливают их поглощение. Например, при снабжении небольшой пряди корней легкодоступной фосфорной кислотой в повышенной концентрации интенсивность поглощения фосфора этой небольшой частью корней резко увеличивается. Это объясняет эффективность локального внесения удобрений, несмотря на то что в контакт с удобрениями входит небольшая часть корней. Изучение функции зародышевых и узловых корней показало важную роль узла кущения в распределении воды и минеральных веществ. Узел кущения состоит из рыхлой пористой паренхимной ткани, поэтому вещества, передвигающиеся по проводящим путям корней, легко переходят из одного проводящего сосуда в другой и, следовательно, могут быть использованы любой надземной частью растений. Корни узла кущения обладают высокой поглотительной способностью и играют более значительную роль в питании растений, чем первичные (зародышевые) корни. Особенно возрастает их роль в период кущения злаков, когда происходит усиленное ветвление узловых корней. Зародышевые корни частично участвуют в питании растений и на более поздних этапах развития. Таким образом, различные зоны корня различаются по способности к поглощению ионов.
Корни являются не только органами поглощения минеральных элементов и воды. Они обладают синтетической способностью. В них образуются многие органические соединения: белки, аминокислоты, амиды, алкалоиды, фитогормоны, в частности цитокинин, и др. (рис. 4.2). Поглощение элементов минерального питания растениями в зависимости от характера затрачиваемой энергии может быть активным и пассивным. Активное поглощение требует затрат метаболической энергии, пассивное же поглощение происходит без затрат метаболической энергии, оно совершается за счет энергии тепловой диффузии или за счет солнечной энергии.
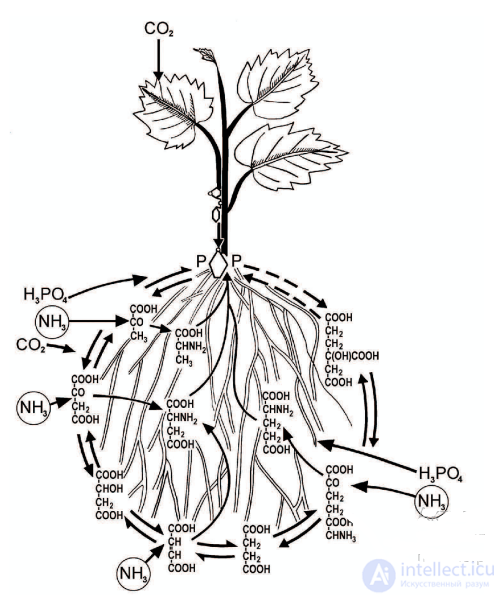
Рис. 4.2. Круговорот веществ и метаболическая роль корней
Активное поглощение и передвижение ионов осуществляются по системе, состоящей из протопластов клеток, связанных между собой тяжами протоплазмы — плазмодесмами. При пассивном передвижении ионы, достигнув поверхности корня, либо посредством массового тока, либо посредством диффузии попадают в свободное пространство корня и далее с транспирационным током передвигаются по растению. Клетки растений (в отличие от животных клеток) имеют рыхлые целлюлозные оболочки, которые, соединяясь между собой, образуют непрерывную систему, так называемый апопласт. По этой системе вследствие транспирации воды листьями и происходит ее движение и растворенных в ней веществ. Ионы в свободном пространстве передвигаются и посредством диффузии. Диффузия всегда происходит от большей концентрации к меньшей. Процесс этот медленный, например краска флуоресцеийн диффундирует за 1 ч Ha 5 мм, за 24 ч — на 25 мм, a за год — на 50 см. Поэтому диффузия не играет роли в передвижении растворимых веществ в растении на большие расстояния, например из корня в лист. Ионы, вошедшие в контакт с корнем, адсорбируются клеточными стенками. Процесс адсорбции ионов корнями носит обменный характер. Высокую интенсивность обмена веществ, значительную скорость поступления и передвижения веществ в растениях можно объяснить адсорбционным обменом между корневой системой растений, с одной стороны, и почвенными коллоидами (твердая фаза), а также почвенным раствором (жидкая фаза) — с другой. Между корневой системой растений и почвенными коллоидами, а также почвенным раствором существует тесный контакт. Корни растений в почве сильно разветвлены. Они проникают на глубину 1,5-2 м, а у отдельных растений — до 5-10 м и более. Например, в засушливые годы корни люцерны иногда проникают на глубину до 18м. В ширину корни культурных растений распространяются на 30 - 65 см. Корни и мельчайшие корешки, корневые волоски густой сетью опутывают почву и ее коллоидные частицы. И если учесть, что корневые волоски живут в течение одних или нескольких суток и что их число огромно (например, на 1 мм? поверхности корня кукурузы имеется 425 корневых волосков и больше, а в среднем у большинства сельскохозяйственных культур — 200-500), то станет ясным совершенный контакт, который существует между почвой и корневой системой растения, ee корневыми волосками, через которые поступает пища. Благодаря этому контакту и происходит процесс обменной адсорбции, сущность которого состоит в следующем. Питательные ионы (например, K*, Са”, Ме”, NH,", МОу, H,PO,, SO) поступают в растения через корневую систему в обмен Ha ионы HY, НСО, расположенные на поверхности корневых волосков и возникающие при дыхании корней. Корневая система растений выделяет большое количество углекислоты, например горчица за 85 дней жизни выделяет 2,25 т СО, с 1 га, то же самое можно сказать и о других сельскохозяйственных культурах. Появляющийся углекислый газ, реагируя с водой, образует углекислоту: CO, + H,0 — H,CO,. Как слабая кислота, она частично диссоциирует на ионы Н’ и НСО,. Так что в результате дыхательного процесса на поверхности корневых волосков постоянно появляются все новые и новые порции ионов Н’ и НСО,. Катионы почвы К’, Са”, Ме”, МН, и другие немедленно вступают в обменную реакцию и вытесняют с поверхности корневого волоска катион HY. Анионы же МОу, H,PO,, SOF и другие обмениваются и вытесняются в раствор анионом НСО}. Появившиеся на поверхности корневого волоска питательные катионы и анионы неизбежно входят в соприкосновение с базоидной (основной) и ацидоийдной (кислотной) частями плазмы клетки, и здесь они дают начало различным органическим соединениям или передвигаются до листьев, где также синтезируются органические вещества. Передача ионов от клетки к клетке происходит по принципу адсорбции-десорбции от одной молекулы белка к другой вплоть до проводящей сосудистой системы корня. Этот физикохимический процесс протекает с большой скоростью. Важно учесть, что поглощение питательных элементов корневой системой растений может происходить не только в обмен на ионы H* u НСОу, образующиеся в процессе дыхания, но и на ионы органических и минеральных соединений, выделяемые корнями. Установлено, например, что корни растений выделяют лимонную, яблочную, щавелевую и другие органические кислоты, которые слабо диссоциируют, но все же распадаются Ha Н’ и органические анионы. Эти ионы находятся на поверхности корневых волосков и могут принимать участие в обменных реакциях на соответствующие катионы и анионы почвенного раствора. При более тесном контакте корневой системы с почвенным поглощающим комплексом поглощение питательных веществ растением протекает более интенсивно. Наилучшие условия питания растений создаются при достаточном количестве ионов как в почвенном растворе, так и в адсорбционно-связанном состоянии. Подтверждением существования адсорбционного поглощения питательных элементов является наличие определенной емкости поглощения катионов и анионов у корневых волосков, так же как и у почвенных коллоидов. Величина емкости поглощения корней зависит от вида растений, условий их питания и других факторов. Например, емкость поглощения катионов на 100 г сухого вещества корней составляла у бобовых культур 40-60 ммоль, у картофеля и томатов — 35-38, у злаков — 9 - 29 ммоль. При питании растений азотом повышается емкость катионного поглощения у корней, что можно объяснить, по-видимому, большим синтезом белковых веществ. Значительную часть емкости катионного поглощения составляют ионы водорода (HY), которые обмениваются на катионы почвенного раствора, необходимые для питания растений. Анионное поглощение менее изучено, но в принципе оно аналогично катионному обмену. Важную роль здесь должен играть анион НСО. У многих культур анионный обмен количественно превышает катионный, что свидетельствует о наличии большего количества коллоидов активной части корней растений с положительным 3aрядом. Таким образом, клетки адсорбируют на своей поверхности положительно или отрицательно заряженные ионы минеральных солей, которые могут обменно вытесняться в свободное пространство другими ионами того же заряда. Основным барьером для поглощения ионов и веществ является поверхностная мембрана, или плазмалемма. Способность растений поглощать или обмениваться ионами со средой в значительной мере зависит от свойств мембран. Отсюда понятны попытки исследователей воздействовать на мембраны в целях регуляции поступления ионов в корни. В этом отношении представляют интерес мембранно-активные соединения, с помощью которых можно воздействовать на ионный транспорт в растениях. К ним относятся антибиотики (валиномицин, грамицидин и др.), 2,4-динитрофенол, диметилсульфоксид и др. Наиболее перспективный из них — диметилсульфоксид, обладающий мягким действием. Он увеличивает проводимость сульфолипидного слоя мембран. Усиление поглощения и передвижения ионов нитратов и фосфатов в растениях сахарной свеклы, обработанных в условиях полевого опыта 2,5-5%-м водным раствором диметилсульфоксида, а также активация оттока ассимилятов из листьев в корни способствовали повышению эффективности использования питательных веществ почвы и удобрений, активировали рост и повышали урожай корнеплодов на 30 - 60 ц/га, увеличивали сбор сахара на 5 - 10 ц/га. Поглощение питательных веществ — это сложный физико-химический и метаболический процесс, включающий диффузию, адсорбцию и метаболический перенос веществ против электрохимического градиента. Диффузия важна при перемещении растворимых питательных веществ в почве к корням растений, а обменная адсорбция — при поступлении питательных элементов в растение через корневую систему. В дальнейшем поступившие питательные элементы взаимодействуют с протоплазмой клетки метаболическим или неметаболическим путем. Метаболическое поглощение и перемещение питательных веществ происходят очень быстро и зависят от аэробного дыхания, температуры и аэрации почвенного раствора. При этом поглощении важную роль играют макроэргические соединения, в частности АТФ, снабжающие энергией этот процесс. Неметаболическое, или пассивное, поглощение непосредственно может быть не связано с жизнедеятельностью растений и поэтому мало зависит от температуры и других условий жизни растительного организма. Примером неметаболического поглощения является пиноцитоз — захват части питательного раствора, при котором клетками молодых корешков могут поглощаться ионы, молекулы, их агрегаты и капельки раствора. Существуют три механизма подачи питательных веществ к поверхности корня: 1) корневой перехват, 2) массовый поток, 3) диффузия. Вклад каждого из этих механизмов зависит от интенсивности поглощения веществ корнем и от обеспеченности почвы питательными веществами. Корневой перехват. Корни в процессе роста движутся в почве, соприкасаясь с питательными веществами, поглощают их. Доля корневого перехвата в питании небольшая, так как объем корневой системы в почве на глубине 15 см не превышает 0,5 - 2% от общего объема почвы. Корневой перехват играет существенную роль при содержании в почве питательных веществ в больших количествах по сравнению с потребностями растения. Если же их меньше, чем это нужно для обеспечения максимальной потребности растений, большая часть питательных веществ, усваиваемых корнями, обеспечивается массовым потоком и диффузией.
Массовый поток. Корни растений поглощают из почвы воду, что вызывает движение почвенного раствора через толщу почвы к корням. Так как в почвенном растворе содержатся питательные вещества, то они и переносятся массовым потоком к поверхности корня, становясь доступными для поглощения. В зависимости от вида растения и погодных условий интенсивность потока воды может сильно изменяться, но обычно она находится в пределах от 1 до 6:10” см° воды на 1 см? корневой поверхности в секунду. Диффузия. Поглощение корнем питательного вещества сопровождается уменьшением его концентрации у поверхности корня и возникновением градиента концентрации. А это делает возможной диффузию питательного вещества к корню. Скорость диффузии ионов через почву изменяется в зависимости от типа почвы и природы поглощения ионов почвой. Для ионов, не адсорбируемых почвой, например нитратов, градиент концентрации (соотношение между ионами в растворе и ионами, адсорбируемыми на поверхности почвенных частиц) может достигать 1. Для сильно поглощаемых почвой ионов, таких, как фосфат, это соотношение может быть 107. Чтобы корень мог поглотить ион нитрата, последний может находиться от него на расстоянии до 1 см и даже более, а для поглощения иона фосфата — не более 0,1 см. Между тем корни однолетних растений часто размещены в почве друг от друга в среднем на расстоянии 0,5 см. Для различных питательных элементов участие указанных механизмов неодинаково. Так, фосфор и калий доставляются к корням в основном путем диффузии, а кальций и магний — посредством массового потока. Массовый поток приобретает важную роль при более высокой концентрации нитратов в почвенном растворе (около 10 моль МО, /л). При низких концентрациях нитрата большое значение имеет диффузия. Концентрация нитратов в прикорневом слое почвы в 3-4 раза выше, чем в почве внекорневого слоя, что особенно заметно при внесении высоких доз азота (120 кг/га). Такое повышение концентрации нитратов в почве, прилегающей к корню, связано с тем, что нитраты передвигались к корню путем массового потока, но при этом скорость поглощения воды корнями (и транспирация) была выше скорости поглощения нитратов. Поскольку и массовый поток, и диффузия в основном зависят от концентрации соответствующего вещества в почвенном растворе, уровень питательных веществ почвенного раствора является фактором, в первую очередь определяющим доступность питательных веществ растению.
Все, что сказано о механизмах подачи питательных веществ к поверхности корня, относится к той их части, которая находится в почвенном растворе и наиболее доступна для непосредственного использования растениями. Растение не является только потребителем уже готовых для него питательных веществ в почве. Корни растений активно воздействуют на почву, находящуюся у их поверхности. Хорошо известна способность корней выделять во внешнюю среду органические и минеральные вещества (сахара, органические кислоты, азотсодержащие органические соединения, витамины, ферменты и др.). Выделенные корнями органические вещества служат пищей для микроорганизмов, которые в процессе жизнедеятельности способствуют мобилизации питательных веществ почвы, повышая их доступность для растений в участках, непосредственно примыкающих к корням. Но значение микроорганизмов состоит не в том, что они снабжают растение органическими формами азота и фосфора, которые по эффективности уступают минеральным. Микроорганизмы снабжают растения физиологически активными веществами (ауксинами, витаминами, антибиотиками), которые в определенных условиях могут даже в ничтожных количествах оказывать положительное влияние на рост растений. Такое действие корневых выделений растений на доступность питательных веществ почвы с помощью ризосферных микроорганизмов является косвенным. Растения оказывают и прямое воздействие на труднодоступные для них соединения почвы (особенно фосфора), переводя их в усвояемые формы. Еще Д.Н. Прянишников доказал способность люпина, гречихи, горчицы использовать фосфор трехзамещенных фосфатов или естественных фосфоритов. Способность этих растений усваивать фосфор труднодоступных фосфатов связывается прежде всего с кислотностью корневых выделений. Так, в растворе, окружающем корневые волоски люпина, рН составляет 4-5, а клевера — 7-8. Роль корневой системы исключительно важна и в синтетических процессах. Корни являются не только органами накопления и передачи из почвы питательных веществ, HO и Opганами синтеза органических веществ. Многочисленные исследование, проведенные в 30-60 годы прошлого столетия показали, что корневой системе растений присущи многие синтетические процессы. При изучение пасоки корней различных растений в ней обнаружен широкий спектр растворимых органических соединений: аминокислоты, олигопептиды, сахара и ростовые вещества. Значительный вклад в изучение физиологии корневой системы, ее проницаемости
и способности поглощать, выделять и перерабатывать минеральные вещества и некоторые органические соединения внес Д.А. Сабинин. Он одним из первых предположил, что корневая система является не только органом поглощения, но и синтеза и превращения веществ. По существу, эти работы стали началом нового направления в изучении минерального питания растений, утвердившего представление о синтетической способности корневой системы. В дальнейшем это направление получило развитие в работах многих исследователей. С помощью стабильного изотопа азота “№ Ф.В. Турчиным (1965) было установлено, что практически весь поглощенный корневой системой аммонийный азот (SNH) и значительная часть нитратного ("№О,) находится в пасоке корня в виде различных азотсодержащих органических соединений. В корнях различных сельскохозяйственных культур синтезируется около 18 аминокислот из 20 входящих в белки растений. Исследования, проведенные с меченым фосфором (*Р), серой и рядом микроэлементов показали, что еще в корнях растений анион фосфорной кислоты (H,PO,) включается с помощью эфирных связей в состав различных органических соединений, а сульфат-ион (SO), восстанавливаясь образует серосодержащие аминокислоты: цистин, цистеин и метионин. Убедительные доказательства синтетической деятельности корневой системы были получены учеником Д.Н. Прянишникова академиком А.А. Шмуком (1941) совместно с сотрудниками. Они установили, что никотин синтезируется корневой системой табака, а не листьями. Если привить табак на томат, то в листьях табака никотина практически не обнаруживается, и, наоборот, в листьях томата привитого на табак, накапливается 3 - 4% никотина. Неоспоримым доказательством синтетической деятельности корневой системы является также быстрое появление новых побегов (поросли) после удаления надземной части у корневищных и корнеотпрысковых, в т.ч. плодовых и ягодных растений. Синтетическая деятельность листа и корня растений тесно связана. Например, с увеличением поступления питательных веществ через корни усиливаются дыхание и приток к ним углеводов из листьев, а из корневой системы в надземную часть до листьев и генеративных органов движутся аминокислоты и другие органические соединения. Открытие разнообразной синтетической деятельности корневой системы — одно из самых крупных достижений науки в ХХ столетии в области физиологии корневого питания растений. Корень — такая
же лаборатория синтеза, как и лист, т.е. биосинтез сложных органических веществ происходит при взаимосвязанной синтетической деятельности листа и корня. Энергия для всех жизненно необходимых превращений и передвижений веществ освобождается при дыхательных процессах, непрерывно совершающихся во всех живых клетках и тканях растений. Во всех этих передвижениях и превращениях видную роль играет белковая плазма клетки, которая имеет двойную природу — базоидную и ацидоидную. В белковой плазме, ee молекулах ацидоидная и базоидная части расположены мозаично, так что при передвижении питательных веществ от клетки к клетке происходят постепенный обмен и взаимодействие катионов и анионов с положительно заряженными (базоидными) и отрицательно заряженными (ацидоидными) частями белковых молекул плазмы клеток. Это свойство живой клетки нужно всегда учитывать, чтобы понять процессы поступления, перемещения и превращения питательных веществ в растительных организмах. Растению присуща избирательная способность: оно поглощает больше тех элементов, в которых нуждается, и меньше тех, которые ему не нужны. Это объясняется физиологическими законами живого организма. Например, при внесении в почву селитры NaNO; растение больше поглощает анион МО, и меньше катион Ма’. При внесении сульфата аммония (NHy),SO, растение больше поглощает катион NH4" и меньше — анион SO и т.д. Вещества, физиологически необходимые растению, по мере поступления через корни тут же подвергаются процессам синтеза и ассимилируются (усваиваются), переходят в другие соединения, в частности органические, т.е. уходят с адсорбционной поверхности корневого волоска. А те вещества, в которых растения не нуждаются, не подвергаются изменению, остаются в нем в минеральной, легкорастворимой ионной форме в том же виде, в каком были до поступления в растение. После выравнивания их концентрации в клеточном и почвенном растворах на поверхности корневого волоска и на поверхности почвенных коллоидных частиц они перестают поступать в растение, так как поступление ионов из почвы происходит по принципу адсорбции-десорбции (адсорбируется столько, сколько и десорбируется, согласно закону действующих масс). Нужные же растению питательные элементы (их анионы и катионы) будут усваиваться благодаря синтезу из них органических соединений, т.е. исчезать с поверхности волосков. И следовательно, равновесие нужных ионов будет постоянно нарушаться до тех пор, пока будут усваиваться эти питательные элементы растением.
Об избирательности поглощения питательных веществ растениями и активном их поступлении из почвенного раствора в корни свидетельствует тот факт, что концентрация солей ряда элементов питания в клеточном соке растений гораздо выше, чем в питательном растворе, в который погружена корневая система. Например, концентрация калия в пасоке кукурузы была в 20 раз, фосфора — в 14, а кальция — в 4 раза выше, чем во внешнем питательном растворе. С завершением жизненного цикла приостанавливается поступление питательных веществ в растения, прекращается использование катионов и анионов и наступает равновесие концентрации их на поверхности корневого волоска и в почвенном растворе, а также на поверхности коллоидных частиц. Так как растения избирательно поглощают ионы (одних больше, других меньше, в зависимости от физиологической потребности в них), то минеральные удобрения (соли) могут быть или физиологически кислыми, или физиологически щелочными. Если растение больше поглощает катионы соли, а анионы ее болыше накапливаются в почвенном растворе, то такая соль будет физиологически кислая. Например, физиологически кислыми являются такие минеральные удобрения, как сульфат аммония, хлористый аммоний, аммиачная селитра, хлористый калий и др. Если же растение больше поглощает анион соли, а катион ее накапливается в почвенном растворе, то такая соль будет физиологически щелочной. К ним относятся натриевая селитра, кальциевая селитра и др. При наличии в почвенном растворе токсичных веществ (тяжелых металлов, пестицидов и т.п.) значительная их часть задерживается уже в корнях, та часть из них, которая все же проникает в стебли и листья задерживается в свою очередь в этих надземных органах и лишь незначительная часть токсикантов поступает в семена. Таким образом, механизмы ограничивающие поступление и накопление токсичных веществ в репродуктивные органы присутствуют не только в корнях, но и в вегетативных органах, что очень важно с точки зрения использования растительной продукции. Поступление питательных веществ в растение происходит избирательно и зависит от интенсивности дыхательных процессов и прежде всего от энергии дыхания корней и выделения ими ионов Н’ и НСО). Важную роль играет степень развития корневой системы, увеличение ее поглощающей поверхности и усвояющей способности. В жизни растения можно различить два периода питания, © KOторыми следует считаться при использовании удобрений.
Первый, получивший название критического, совпадает с начальными фазами роста и развития большинства растений. В этот период растения особо чувствительны как к недостатку, так и к избытку питательных веществ. Химический состав растений в начальные фазы роста характеризуется высоким содержанием азота и многих зольных элементов. В этот период растения предъявляют повышенные требования к условиям минерального питания. Второй период получил название периода максимального потребления питательных веществ. Он характерен для более поздних фаз развития и определяется биологическими особенностями растений. Поступление питательных веществ в злаковые растения, за исключением кукурузы, к концу колошения почти заканчивается, хотя к этому времени они образуют не более 50-60% растительной массы OT полного урожая (табл. 4.2). Так, озимая пшеница при хорошем развитии уже в осенний период усваивает азота и калия 43 - 47%, в то время как сухая масса растений составляет не больше 10% полного урожая. Это полностью относится и к озимой ржи, которая за осенний период усваивает до 50 - 60% азота, фосфора и калия. Ячмень и овес уже в фазу цветения поглощают 100% калия, а после происходит даже потеря этого питательного элемента (экзоосмос, или выделение). Накопление питательных веществ кукурузой происходит медленнее. Даже к началу цветения поступает только 30-40% азота и калия и 15% фосфора от содержания этих элементов в кукурузе при ее созревании. Сахарная свекла, картофель, капуста и другие овощные культуры отличаются более продолжительным или растянутым периодом питания. Азот, фосфор и калий они усваивают почти в течение всего вегетационного периода (табл. 4.3).
4.2. Динамика накопления питательных элементов в растениях, % от максимума
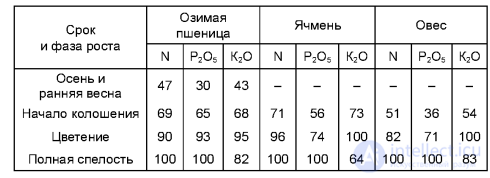
4.3. Динамика поступления питательных элементов в растениях сахарной свеклы (данные Научно-исследовательского института сахарной свеклы), % от максимума
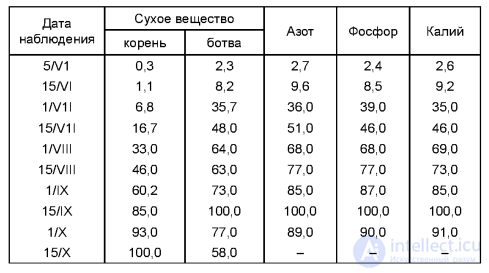
Таким образом, период питания не равен периоду вегетации растений. У многих растений он значительно короче периода вегетации (конопля и большинство злаковых культур). У других же культур он растянут и почти совпадает с периодом вегетации растений (сахарная свекла, картофель, капуста и другие овощные культуры). Итак, питание растений с учетом их биологических особенностей можно регулировать по периодам роста, что позволяет формировать величину и качество урожая. Периодичность питания растений является теоретическим обоснованием дробного внесения удобрений (в разные сроки и в разные слои почвы). Внесением удобрений в один прием и в один слой почвы не всегда можно добиться полного использования их потенциальных возможностей. Дозы легкорастворимых минеральных удобрений, достаточные для критического периода питания, будут малы для периода максимального потребления питательных веществ. И наоборот, большая доза вредна для первого (критического) периода, когда молодые корешки растения чувствительны к высокой концентрации питательных веществ. Вот почему правильная система питания растений предусматривает сочетание основного (на глубину 18-25 см), припосевного удобрения (на глубину 5-8 см) и подкормок в период роста растений (на глубину 10 - 15 см).
Исследование, описанное в статье про корневое питание растений , подчеркивает ее значимость в современном мире. Надеюсь, что теперь ты понял что такое корневое питание растений и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Агрохимия и биохимия
Ответы на вопросы для самопроверки пишите в комментариях, мы проверим, или же задавайте свой вопрос по данной теме.
Комментарии
Оставить комментарий
Агрохимия и биохимия
Термины: Агрохимия и биохимия