Лекция
Привет, Вы узнаете о том , что такое мембрана нейрона , Разберем основные их виды и особенности использования. Еще будет много подробных примеров и описаний. Для того чтобы лучше понимать что такое мембрана нейрона , настоятельно рекомендую прочитать все из категории Нейробионика.
Мембрана содержит пассивные и активные участки.
Пассивная мембрана обладает способностью проводить
отклонения потенциала в какой-либо точке от равновесного
значения к другим точкам с затуханием, пропорциональным
расстоянию. В активной мембране распространяется
волна регенерирующего, возбуждения за счет
вольт-зависимого изменения проводимости ионных каналов.
В спокойном состоянии мембраны среда внутри клетки
имеет некоторый «потенциал покоя» (примерно 70 мВ)
относительно внешней среды. Эта разность потенциалов
определяется разной концентрацией ионов ряда элементов,
главным образом ионов натрия и калия. В покое
концентрация ионов калия на порядок ниже, чем с внутренней
стороны (внутри нервной клетки или волокна).
Внутренняя поверхность мембраны заряжена
отрицательно по отношению к ее наружной поверхности.
Наблюдаемая разность концентраций ионов и потенциалов по обе
стороны мембраны обусловлена процессами клеточного
обмена — неким гипотетическим «метаболическим
насосом», выкачивающим часть ионов натрия наружу и
накачивающим часть ионов калия внутрь клетки.
Внутриклеточный потенциал может повышаться
(деполяризация) или еще более понижаться (гиперполяризация).
Изменение потенциала в сторону деполяризации называют
также возбуждающим постсинаптическим потенциалом
(ВПСП), а в сторону гиперполяризации — тормозным
постсинаптическим потенциалом (ТПСП). Соответственно
все факторы, включая действие синапсов, называют
возбуждающими или тормозными в зависимости от того,
повышают или понижают они потенциал с внутренней
стороны мембраны.
Возбуждающее воздействие, вызывающее повышение
внутриклеточного потенциала (деполяризацию), не
приведет к активному возбуждению мембраны, если не
достигнут уровень возбуждения, т. е. изменение потенциала было
меньше порогового. При этом порог по потенциалу зависит
от формы возбуждающего импульса: чем положе
восходящий фронт импульса, тем выше порог. Подпороговые
изменения потенциала сопровождаются трансмембранным
током, образованным емкостным током и током утечки.
При подпороговых процессах проницаемость мембраны
для ионов существенно не меняется, поэтому и процессы
относят к пассивным.
Активный процесс начинается с того момента, когда
внутриклеточный потенциал превысил уровень порогового
возбуждения. С этого момента резко увеличивается
проницаемость мембраны для ионов натрия. Проходя
снаружи через мембрану внутрь клетки (волокна), ионы натрия
еще больше повышают внутренний потенциал, повышение
потенциала увеличивает проницаемость для ионов
натрия. Процесс нарастает лавинообразно до тех пор, пока
разность потенциалов между внутренней и внешней
средами не достигнет противоположного знака, затем ток
натрия убывает. Зато постепенно увеличивается
проницаемость мембраны для ионов калия. Калиевый ток течет
навстречу натриевому. Он также достигает максимума и
затем спадает. За счет тока натрия этот потенциал
достигает своего максимума, а ток калия вновь сводит
потенциал мембраны к значению, свойственному потенциалу
покоя.
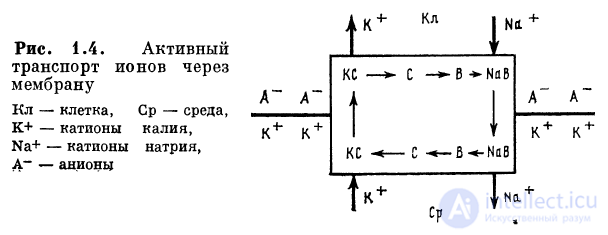
1.2.1. Активный транспорт ионов через мембрану
По современным представлениям в нейроне исходно
существуют различные концентрации ионов калия внутри
и снаружи клетки. Потенциал равновесия «запирает»
выход ионов калия из клетки. Однако отрицательный
потенциал создает условия движения в клетку ионов натрия
из внешней среды, что в свою очередь снижает уровень
отрицательного мембранного потенциала, облегчая выход
калия под влиянием разницы концентраций. Постепенно
концентрации ионов калия и натрия внутри и снаружи
будут выравниваться. Для того чтобы поддержать эту
разницу концентраций, должен существовать механизм вывода
ионов натрия из клетки и переноса ионов калия в клетку.
Этот механизм называют натрий-калиевым насосом.
Внутриклеточный натрий на внутренней стороне мембраны
соединяется с веществом — носителем В. Комплекс NaB
электронейтрален и благодаря диффузии выходит наружу,
где расщепляется. Освободившийся носитель В под
влиянием энзима переходит в форму С и присоединяет ион
калия К+. Комплекс КС диффундирует внутрь клетки и
отдает ион калия. Переносчик С при участии АТФ переходит
в форму В с использованием внутренней энергии клетки
(рис. 1.4), и цикл повторяется.
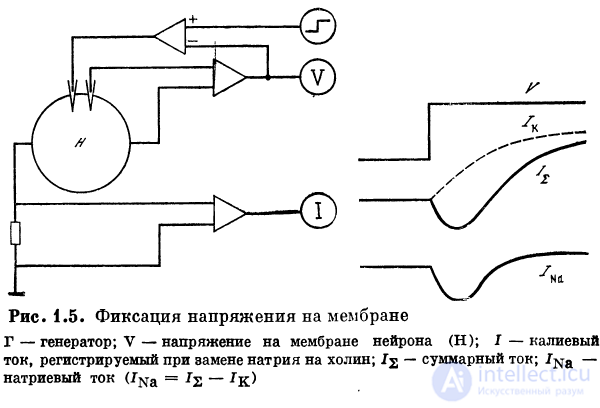
1.2.2. Элекгротонический потенциал. Фиксация напряжения
При подаче импульса тока через введенный в тело нейрона
микроэлектрод можно обнаружить, что потенциал
нейрона, регистрируемый вторым микроэлектродом,
изменяется. Нейрон деполяризуется при инъекции
положительных зарядов д гиперполяризуется при введении
отрицательных. Схематически мембрану нейрона можно
представить как параллельно подключенные емкость и
сопротивление. Емкость образована диэлектриком — фос-
фолипидами мембраны. Сопротивление создают каналы
ионной проводимости. При достижении мембраной потен-циала покоя, определяющегося соотношением
потенциалов равновесия разных ионов, суммарный ток равен нулю. Об этом говорит сайт https://intellect.icu .
При подаче импульса тока сначала он течет через
емкость (емкостный ток). Падение напряжения на
сопротивлении при этом равно нулю. По мере зарядки
конденсатора все большая доля тока течет через сопротивление.
После зарядки конденсатора ток течет через мембрану,
создавая на ней падение напряжения (рис. 1.5).
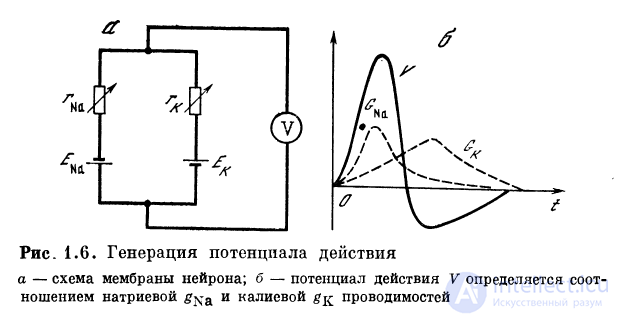
1.2.3. Потенциал действия
Методом фиксации напряжения было показано, что
мембрана нейрона имеет быстро инактивирующуюся
натриевую проводимость и устойчиво сохраняющуюся калие-
вую. Если не учитывать влияние емкостей, то нейрон
можно представить схемой, состоящей из двух батарей и двух
переменных сопротивлений. Одна батарея определяется
потенциалом равновесия для калия (—75 мВ). Переменное
сопротивление воспроизводит изменение калиевой
проводимости. Другая батарея определяется потенциалом
равновесия по натрию (+55 мВ) и переменным
сопротивлением, воспроизводящим натриевую проводимость. Эти
батареи и сопротивления образуют цепь, формирующую
потенциал действия. При этом падение напряжения на
мембране определяется соотношением проводимостей
(рис. 1.6).
1.2.4. Суммация электротонических потенциалов
Электрические свойства мембраны нейрона
характеризуются двумя константами: постоянной времени и
постоянной длины. Постоянная времени мембраны определяет
временную суммацию электротонических потенциалов:
при поступлении на один и тот же участок мембраны двух
электротонических потенциалов они суммируются на
интервале времени, пропорциональном постоянной времени.
Постоянная длины мембраны определяет
пространственную суммацию электротонических потенциалов: если два
электротонических потенциала одновременно поступают
на два соседних участка пассивной мембраны, то к
собственному потенциалу каждого участка добавляется часть
потенциала соседнего участка, пропорциональная
расстоянию между участками, деленному на постоянную длины
мембраны.
Когда электротонические потенциалы возникают в
разных участках мембраны в разные моменты времени, то их
суммация определяется как постоянной времени, так и
постоянной длины мембраны — имеет место так
называемая пространственно-временная суммация.
Клетки с большими ^значениями этих констант более
эффективно суммируют сигналы. Суммация позволяет
мембранному потенциалу достичь критического значения
генерации потенциала действия даже тогда, когда
отдельные электротонические потенциалы этого уровня не
достигают. Электротонические потенциалы определяются
пассивными свойствами мембраны — емкостью и
сопротивлением. Однако источником электротонических
потенциалов в пассивных участках мембраны могут быть
происходящие в прилежащих участках изменения
потенциала, вызванные изменением ионной проводимости. Так,
потенциал действия, генерируемый вследствие
открывания натриевых каналов, создает в прилегающих участках,
еще не охваченных процессом открытия натриевых
каналов, деполяризационный сдвиг, определяемый константой
длины мембраны. Аналогичным образом постоянная
времени и постоянная длины вносят свой вклад в суммацию
электротонических потенциалов, вызванных другими
источниками: постсинаптическими потенциалами и пейсме-
керными потенциалами.
1.2.5. Пейсмекерные потенциалы
В ряде нейронов даже после полной изоляции их сомы
(ампутации дендритов и аксонов) при внутриклеточной
регистрации наблюдаются колебания потенциала
амплитудой 10—20 мВ и частотой от 1,0 до 2,5 Гц, получившие
название пейсмекерных потенциалов. Эти потенциалы
могут быть регулярными или образовывать отдельные
веретена, в которых амплитуда потенциалов постепенно
возрастает, достигает максимума и падает. Сравнение
пейсмекерных потенциалов нейрона до и после изоляции от
других нервных клеток указывает на то, что источником
колебаний являются эндогенные процессы, поскольку тип
активности сохраняется и в полностью изолированном
нейроне. Если пейсмекерные потенциалы достигают
порога генерации потенциала действия, то они запускают их
генерацию.
Таким образом, благодаря наличию пейсмекерных
потенциалов нейрон становится активным генератором.
Более того, поскольку определенный тип нейрона и при
полной изоляции сохраняет свой тип пейсмекерной активности,
эта активность является генетически заданной. Вместе
с тем пейсмекерный потенциал очень чувствителен к
изменениям уровня мембранного потенциала: небольшая
деполяризация клетки током до 1 нА достаточна для того,
чтобы увеличить частоту и амплитуду пейсмекерных
потенциалов, а гиперполяризация в тех же пределах ведет к
замедлению пейсмекерных колебаний, их урежению,
снижению амплитуды и, наконец, полному подавлению.
Анализируя пейсмекерные потенциалы, можно заметить, что при
возрастании их амплитуды деполяризационная волна
становится более острой. Это объясняется тем, что к пейсме-
керному потенциалу присоединяется локальный
потенциал, не достигающий порога развития лавинообразного
процесса, связанного с генерацией потенциала действия.
Чувствительность пейсмекерных потенциалов к сдвигам
мембранного потенциала делает их легко управляемыми
внешними влияниями.
У ряда нейронов можно обнаружить несколько локу-
сов пейсмекерных колебаний с разными свойствами и
разным вкладом в механизмы генерации потенциалов
действия. Источником пейсмекерных колебаний, по-видимому,
являются электрогенные эффекты активного транспорта
ионов. Переходы от деполяризации к гиперполяризации,
образующие пейсмекерную волну, не сопровождаются
изменением сопротивления мембраны. Активный транспорт
ионов натрия ограничивается при развитии
гиперполяризации. Активный транспорт ионов хлора ограничивается
развитием деполяризации. Таким образом,
пейсмекерный потенциал имеет зону оптимума, ограниченную
сверху определенной деполяризацией, а снизу — определенной
гиперполяризацией нейрона.
1.2.6. Электрический синапс
Электрический синапс образован срастанием мембран пре-
синаптического и постсинаптического нейронов. Ток,
инъецируемый в один нейрон через каналы, связывающие
обе мембраны, затекает в постсинаптический нейрон
пассивно и регистрируется на его мембране в виде
электротонического потенциала.
Если один (прбсинаптический) нейрон генерирует
потенциал действия, то у второго (постсинаптического)
обнаруживается электротонический потенциал, в
искаженной форме воспроизводящий за счет пассивных свойств
мембраны потенциал действия пресинаптической
мембраны в виде дифазного потенциала. При суммации дифазных
потенциалов постсинаптическии нейрон сам генерирует
потенциал действия, распространяющийся по аксону.
1.2.7. Квантовый механизм химического синапса
Электрический синапс содержит каналы, непосредственно
связывающие пре- и постсинаптическую мембраны и
обеспечивающие электротоническую связь между нейронами.
В отличие от электрического в химическом синапсе
пре- и постсинаптическая мембраны разделены, и
электротонический потенциал не принимает участия в связи
нейронов. Связь реализуется посредством химических
передатчиков — медиаторов. Медиатор находится в
специальных гранулах — везикулах, находящихся в
пресинаптической терминали. Под действием прибывшего в
пресинаптическую терминаль импульса происходит
присоединение везикулы к пресинаптической мембране, и в си-
наптическую щель в сторону постсинаптической мембраны
выбрасывается порция — квант медиатора. При этом
медиатор действует на рецептивный белок
постсинаптической мембраны. Этот рецептивный белок управляет
каналами ионной проводимости. В зависимости от того,
в отношении каких ионов в постсинаптической клетке
регистрируется миниатюрный постсинаптическии потенциал
(0,5—1 мВ) и как изменяется проводимость, спонтанно
выделяются кванты медиатора, образуя в постсинаптичес-
ком нейроне последовательность миниатюрных
потенциалов постоянной амплитуды со случайными интервалами
между ними. Миниатюрные потенциалы исчезают, если
будут блокированы рецепторы, специфичные в отношении
данного медиатора. В регуляции выделения медиатора
принимает участие кальций. При деполяризации кальций
входит в пресинаптическую часть синапса и перемещается
к участкам, где везикулы присоединяются к участкам
пресинаптической мембраны. Кальций облегчает слияние
мембраны везикулы с мембраной терминали, в результате
чего происходит выделение кванта медиатора.
Поступающий к пресинаптическому окончанию
потенциал действия деполяризует термина ли, что приводит
к открытию каналов кальциевой проводимости, которая
в сильной степени зависит от уровня мембранного
потенциала. Поступление кальция приводит к массированному
выходу медиатора сразу из большого числа синаптических
пузырьков. Суммация нескольких выделившихся в ответ
на один потенциал действия квантов медиатора образует
элементарный постсинаптический потенциал.
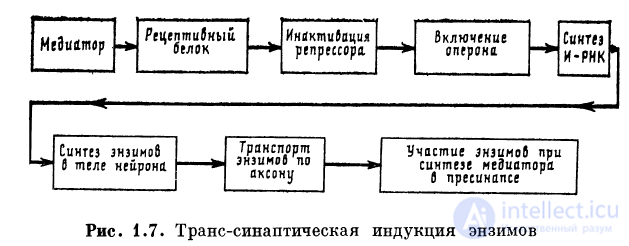
1.2.8. Транссинаптическая индукция энзимов
При действии медиатора на рецептор возникает комплекс,
в котором белок обладает специфической конфигурацией.
Такой белок может выполнять функцию индуктора: транс-
портируясь к ядру клетки и связываясь с репрессором, он
включает механизм транскрипции в определенном оперо-
не, обеспечивая синтез группы функционально связанных
энзимов. Энзимы, поступая к участкам синтеза медиатора,
усиливают в качестве катализаторов синтез медиатора
в пресинаптической терминали нейрона. Таким образом,
реализуется схема медиатор ->- трансформация
рецептивного белка ->- включение оперона на молекуле ДНК -*■
->- транскрипция как синтез и-РНК ->- синтез энзимов ->
->- транспорт энзимов по аксону к пресинаптическому
окончанию -*• усиление синтеза медиатора в пресинапсе
(рис. 1.7). Такой процесс носит название транссинапти-
ческой индукции энзимов.
Наиболее подробно транссинаптическая индукция
энзимов изучена на адренергических нейронах шейного
узла, синтезирующих в термина лях своих аксонов медиатор
норадреналин (или иначе, норэпинефрин). Нейроны этого
узла, обладая мускориновыми и никотиновыми
рецепторами, чувствительны к ацетилхолину. При присоединении
ацетилхолина к никотиновым рецепторам через сутки
в нейроне увеличивается содержание ферментов,
выполняющих функцию катализаторов при синтезе норэпине-
фрина. Синтезируемые в теле нейрона энзимы
транспортируются по аксону к пресинаптическим окончаниям, где
происходит повышенный синтез норэпинефрина.
Вызванное действием ацетилхолина на никотиновые
рецепторы усиление синтеза энзимов опосредовано
процессом транскрипции с участием и-РНК. Это
доказывается тем, что антибиотик актиномицин-D, препятствующий
образованию молекулы и-РНК на матрице ДНК, включает
транссинаптическую индукцию энзимов, катализирующих
синтез норэпинефрина. Следует подчеркнуть, что
индукция энзимов, ответственных за синтез норэпинефрина,
происходит только в том случае, если ацетилхолин действует
на никотиновые рецепторы. Если эти рецепторы
блокировать, то индукции ответственных за синтез норэпинефрина
энзимов не происходит.
Таким образом, аналогичный ферментативной адаптации
механизм включен в регуляцию эффективности работы
нейрона в звене синтеза медиатора.
Анализ данных, представленных в статье про мембрана нейрона , подтверждает эффективность применения современных технологий для обеспечения инновационного развития и улучшения качества жизни в различных сферах. Надеюсь, что теперь ты понял что такое мембрана нейрона и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Нейробионика
Комментарии
Оставить комментарий
Нейробионика
Термины: Нейробионика