Лекция
Это продолжение увлекательной статьи про система из нейроподобных элементов .
...
компоненты
которого будут изменены пропорционально достигнутому
при адаптации уровню чувствительности вторичных
детекторов. В рассматриваемом случае, когда компоненты
F адаптирующего стимула были равны, F не изменит
своего положения и максимум возбуждения останется на том
же вторичном детекторе, который был бы возбужден и
при отсутствии адаптации. Восприятие сигнала после
адаптации к стимулу, порождающему F с равными
компонентами, не изменится. Рели Ft Ф F2, то при адаптации они
изменятся в разной степени, что приведет к изменению
восприятия этого адаптирующего сигнала. Если за
адаптирующим стимулом следует тест-стимул, то адаптация
вызовет компоненты вектора возбуждения, измененные
пропорционально изменению чувствительности каждого
из первичных детекторов. В результате компоненты
вектора возбуждения тест-стимула изменятся в разной степени,
и вектор возбуждения займет иное положение. Максимум
возбуждения на наборе вторичных детекторов также
изменит свое место, и сигнал будет воспринят иначе. Этот
эффект последействия является последовательным
контрастом в данной системе признака, выделяемого набором
детекторов. Вектор возбуждения, создаваемого тестовым
стимулом, будет поворачиваться в направлении от
вектора возбуждения, соответствующего адаптирующему
стимулу.
Эффект адаптации в форме нормализации и
последовательного контраста может быть использован для
расшифровки того, какие именно векторы возбуждения
генерируют разные сигналы. Тот стимул, для которого эффект
нормализации и эффект последействия отсутствуют,
имеет равные компоненты вектора возбуждения. Те стимулы,
которые дают максимальный эффект последействия даже
тогда, когда эффект нормализации не обнаруживается,
образуют ортогональную систему координат. Все другие
сигналы генерируют векторы возбуждения, занимающие
промежуточные значения. Исследование эффектов
адаптации позволяет упорядочить расположение стимулов на
квазирецептивной поверхности, выделив стимулы,
представляющие ортогональную систему координат. Стимулы,
компоненты вектора возбуждения которых равны,
расположены между ними на равном расстоянии. Таким образом, объединяя изучение субъективных различий с
выделением сигналов на основе свойств адаптации, можно
не только выяснить размерность и координаты
перцептивного пространства, но и задать фиксированную систему
координат.
Формальное описание последовательного контраста
основано на матричных преобразованиях с
использованием диагональной матрицы адаптации Л, компоненты
которой равны 1 — yfi, где ft — компонент вектора Ftl
вызванного адаптирующим стимулом St. Вектор возбуждения
F*, появляющийся вследствие последовательного
контраста, будет равен AFj компоненты этого вектора
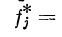
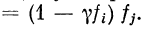
Различительной чувствительностью локального
анализатора называется величина, обратная разностному порогу.
Разностный порог сферической модели восприятия
сигналов характеризуется углом, разделяющим соседние
детекторы. Величина порога на сфере определяется
плотностью распределения детекторов и в процессе адаптации
не меняется. В зависимости от адаптирующего стимула
в процессе адаптации меняется чувствительность
первичных детекторов. При действии мало отличающегося от
адаптирующего тест-стимула вектор возбуждения
поворачивается на угол, характеризующий разностную
чувствительность анализатора. Чувствительность максимальна на
углах в 0° и 90°. Скорость приращения вектора
возбуждения (получаемая дифференцированием вектора
возбуждения, подвергнутого действию оператора адаптации)
характеризует дифференциальную чувствительность.
Максимум дифференциальной чувствительности приходится
на 0° и 90°, а минимум — на 45°.
Выше был рассмотрен локальный анализатор, первичные
и вторичные детекторы которого представляют один
участок рецептивной поверхности. Такие локальные
анализаторы могут перекрывать всю рецептивную поверхность.
В этом случае уже нельзя ограничиваться
исследованием только отдельных локальных анализаторов, поскольку
одноименные первичные детекторы локальных
анализаторов связаны между собой латеральным торможением,
величина которого обратно пропорциональна расстоянию
между рецептивной поверхностью и локальным
анализатором.
Под влиянием латерального торможения одноименных
детекторов векторы возбуждения в каждом из локальных
анализаторов меняются. Эти изменения тем больше, чем
больше различие векторов возбуждения. Если на два
локальных анализатора действуют два одинаковых
сигнала, то направление каждого вектора возбуждения под
влиянием латерального торможения не меняется и
восприятие стимула остается оез изменения. В том случае,
когда на два локальных анализатора действуют два
разных сигнала, вызывающих различные векторы
возбуждения, латеральное торможение одноименных первичных
детекторов приводит к изменению соотношений
компонентов возбуждения. Векторы возбуждения меняют свое
направление, в результате чего стимулы, действующие
на локальные анализаторы, воспринимаются иначе.
Обобщая, можно сказать, что векторы возбуждений
каждого локального анализатора расходятся, угол между
ними увеличивается, а это значит, что субъективное
различие между стимулами увеличивается. Это увеличение
субъективного расстояния между стимулами тем больше,
чем больше исходное различие между векторами
возбуждения и чем ближе друг к другу локальные
анализаторы.
Таким образом, в результате латерального торможения
между одноименными первичными детекторами двух
локальных анализаторов субъективное различие между
стимулами увеличивается. Это явление известно в физиологии
как одновременный контраст.
Обычно имеется целое иоле локальных анализаторов,
одноименные первичные детекторы которых связаны
латеральным торможением. Такое поле обладает
свойством подавления помехи в виде общего для всего множества
анализаторов воздействия. Сигнал же, попадающий на
локальный анализатор, не подавляется. В этом случае
помеха, действуя на все анализаторы, подвергается
сильному латеральному торможению в данном локальном
анализаторе. 4то касается сигнала, то он, действуя
только на локальный анализатор, слабо подавляется помехой.
В результате в локальном анализаторе доля полезного
сигнала возрастает. Чем больше сходство векторов
возбуждения в локальных анализаторах, тем сильнее эффект
торможения. Если сигналы, действующие на тест-поле
и индуцирующее поле, одинаковы, то индуцирующее поле
в той или иной степени маскирует сигнал,
представленный в тест-поле. Маскирующий эффект отсутствует, если
векторы возбуждения ортогональны. Усиление одного
стимула другим возникает тогда, когда векторы в тест-поле и индуцирующем поле противоположны. Чем больше
один вектор другого по модулю, тем больше различие
векторов возбуждения, изменившихся под влиянием латерального торможения. Поскольку латеральное
торможение действует некоторое время после исчезновения
возбуждений, образующих векторы возоуждения,
последующий стимул подвергается действию этого следового
эффекта торможения. ii зависимости от следового эффекта
векторы возбуждения, вызванные действием
последующего стимула, трансформируются. Так, если под
действием стимула вызвано возбуждение в индуцирующем
поле, то торможение, возникшее под его влиянием в тест-поле, ослабит действие такого стимула в области тест-поля. При значительной силе торможения и слабом
входном сигнале возбуждение в тест-поле может быть
полностью подавлено.
Таким образом, на основе механизма латерального
торможения могут быть объяснены явления
одновременной и последовательной маскировки.
Если вектор возбуждения, генерируемый детекторами
одного анализатора, используется для изменения
коэффициентов связей другого анализатора, то он называется
модифицирующим вектором. Модифицирующие векторы
могут генерироваться как первичными, так и
вторичными детекторами. Число компонентов вектора
возбуждения определяется числом детекторов. Компоненты
модифицирующего вектора могут приобретать значения от
нуля до единицы. В том случае, когда компоненты вектора
приобретают только два значения: «нуль» и «единица»,
модифицирующий вектор выполняет переключательную
функцию.
Ранее рассматривался детектор, связи которого были
фиксированными, а поэтому сам детектор был селективно
настроен на определенное значение параметра внешнего
сигнала. Кроме того, следует рассмотреть детекторы,
связи которых с первичными детекторами или
предшествующими селективными детекторами изменяются под
влиянием сигналов, поступающих от других детекторов.
Рассмотрим вторичный селективный детектор,
связанный с первичными детекторами. Коэффициенты этих связей исходно все равны единице. Но коэффициент связи
изменяется (умножается) на возбуждение, управляющее
этой связью. Когда на вектор связи данного детектора
действует модифицирующий вектор, коэффициенты связи
приобретают значения, равные соответствующим
компонентам вектора возбуждения. Коэффициенты связи
остаются модифицированными только до тех пор, пока на
них действует модифицирующий вектор. Отключение
модифицирующего вектора эквивалентно его обращению
в нуль. Поэтому все связи детектора становятся равными
нулю и детектор отключается. Появление нового
модифицирующего вектора устанавливает значения
коэффициентов связи равными или пропорциональными
компонентам вектора возбуждения.
Детектор с вектором связи, изменяемым
модифицирующим вектором, может быть селективно настроен на
любой внешний сигнал в пределах детектируемого
параметра. Для этого нужно, чтобы модифицирующий вектор
соответствующим образом изменил вектор связи. Набор
детекторов, генерирующих модифицирующий вектор,
который управляет коэффициентами связей, назовем
дополнительным набором детекторов.
Если внешний сигнал, генерирующий вектор
возбуждения, меняется одновременно с изменением вектора
возбуждения на дополнительном наборе детекторов, то
детектор с переменными связями все время автоматически
сохраняет максимум возбуждения и становится
инвариантным детектором. Таким образом, инвариантный детектор
может быть селективно настроен на любой стимул в
пределах детектируемого параметра. Это достигается
подачей модифицирующего вектора, который не дает вклада
в основное возбуждение, а только изменяет вектор
связей. Инвариантный детектор обеспечивает автоматический
учет параллельного изменения сигналов в двух
анализаторах. В результате этого на наборе инвариантных
детекторов отображается инвариантное соотношение этих
двух сигналов. Построение инвариантного отображения
приводит к возможности возникновения иллюзий. Так,
если основной сигнал остается постоянным, а меняется
только модифицирующий вектор, то в наборе
инвариантных детекто ров максимум возбуждения будет перемещаться
с одного детектора на другой, создавая иллюзию
перемещения сигнала. Если вектор возбуждения основного
набора детекторов меняется, а модифицирующий вектор не
меняется, то максимум возбуждения перемещается по
набору инвариантных детекторов, адекватно отображая
изменение основного сигнала. Если вектор возбуждения
основного набора детекторов изменяется вместе с модифицирующим
вектором, то максимум возбуждения остается на одном
инвариантном детекторе, адекватно отражая то свойство
объекта, которое связано с соотношением сигналов двух
анализаторэв. Аналогичные рассуждения справедливы
для двух наборов вторичных детекторов. Разница лишь
в том, что компонентами модифицирующего вектора
являются здесь возбуждения вторичных детекторов
дополнительного набора, а вектор связи образован связями
вторичных детекторов с инвариантными детекторами.
Важным типэм модифицирующего вектора является
вектор, все компоненты которого равны нулю, кроме
одного, равного единице. Такой модифицирующий вектор
является переключательным, он подключает к
инвариантному детектору только один вторичный детектор,
устанавливая все коэффициенты связей равными нулю,
кроме одного, равного единице.
Система детекторов с модифицируемыми коэффициентами
связей представляет собой набор инвариантных
детекторов. В простейгпем случае такой набор представляется
одномерным массивом. В более сложных случаях
детекторы могут образовать двумерный массив, образующий
инвариантный экран.
Так как на инвариантном экране производится
константное отображение сигнала, его называют
константным экраном. Двумерный массив детекторов, с которых
информация поступает на константный экран,
представляет собой так называемый аконстантный экран,
формирующий матрицу возбуждений. Каждый элемент аконстантного экрана связан со всеми инвариантными детекторами
единичной матрицей связей. Эта матрица может
изменяться под влиянием модифицирующей матрицы. В
частном случае, когда все элементы модифицирующей матрицы
равны нулю, кроме одного, равного единице, матрица
является переключающей, она выбирает связь данного
элемента аконстаптного экрана с одним из элементов
инвариантного экрана. Аналогичным образолг происходит
подключение всех других элементов аконстаптного экрана. Соответствие между элементами аконстантного и
константного экранов устанавливается при помощи набора
модифицирующих или переключающих матриц. При
постоянстве возбуждения на аконстантном экране изменение
модифицирующих матриц перемещает изображение по
константному экрану, создавая иллюзию движения. При
соответствующем изменении матриц возбуждения и связи
изображение, перемещающееся по аконстантному экрану,
остается неизменным на инвариантном (константном)
экране.
Принцип кодирования номером нейрона относится как
к сенсорным, так и к исполнительным функциям. Если
набор восходящих связей делает детектор селективным
в отношении определенного сигнала, то набор нисходящих
связей превращает единичное возбуждение нейрона в
вектор возбуждения, который порождает специфическую
реакцию. Командный нейрон — это нейрон, возбуждение
которого необходимо и достаточно, чтобы запустить
целостную реакцию или фрагмент целостной реакции.
Командные нейроны можно разделить на первичные,
запускающие целостную реакцию, и вторичные, реализующие
отдельные фрагменты этой реакции. Вторичные
командные нейроны подключены к элементам, выполняющим
функции мотонейронов. Единичный вектор возбуждения
первичного командного нейрона умножается на вектор
связи с вторичными командными нейронами, порождая
новый вектор возбуждения с компонентами, равными
возбуждению вторичных командных нейронов. Компоненты
этого вектора поступают на аналоги мотонейронов и
порождают новый вектор возбуждения, являющийся
управляющим вектором. Совокупность реакций
исполнительных элементов образует специфическую реакцию,
определяющую элемент поведения.
Вектор возбуждения, генерируемый командным
нейроном, будем называть управляющим. К командному
нейрону подключены детекторы. Связи между детекторами
и командными нейронами могут быть жесткими и не
меняться в ходе опыта или же быть пластичными,
изменяться в процессе обучения. К командному нейрону
может быть жестко подключено ограниченное число
детекторов. В этом случае командный нейрон функционирует по принципу «ключ-замок», определяя
селективность реакции. К командному нейрону может быть
присоединено пластическими связями множество детекторов,
в результате чего реакция, запускаемая командным
нейроном, может быть вызвана широким классом сигналов.
В результате изменения коэффициентов связи между
детекторами и данным командным нейроном командный
нейрон может трансформироваться в селективный
приемник сигналов, управляющий реакцией. Реакция
командного нейрона определяется скалярным произведением
вектора возбуждений детекторов предыдущего уровня на
вектор связей этого уровня с командным нейроном.
Управляющий вектор возбуждения, генерируемый первичными
командными нейронами, равен произведению скалярной
величины его возбуждения на вектор связей с вторичными
командными нейронами.
Каждый вторичный командный нейрон также
генерирует управляющий вектор, равный произведению
скалярной величины его возбуждения на вектор связей с
аналогами мотонейронов. Общий управляющий вектор на
уровне аналогов мотонейронов равен сумме управляющих
векторов, порожденных каждым вторичным командным
нейроном.
Управление разными реакциями достигается набором
командных нейронов, каждый из которых генерирует
специфическую реакцию. При помощи латерального
торможения между командными нейронами осуществляется
выбор одной реакции из множества.
Управляющий вектор формируется последовательным
усложнением в каждом слое командных нейронов.
Возбуждение первичного командного нейрона передается по
множеству каналов, формируя состав управляющего
вектора за счет соответствующей системы связей его с
командными нейронами следующего уровня (вторичными
командными нейронами). У некоторых командных
нейронов эти каналы являются пластичными. Отдельные ветви
командного нейрона могут включаться или отключаться,
определяя управляющий вектор. Кроме того, могут
изменяться коэффициенты связей между первичными и
вторичными командными нейронами, изменяя компоненты
управляющего вектора. Аналогичные процессы имеют место в звене вторичный детектор — аналог мотонейрона,
а также в звене мотонейрон — эффектор. Пластичность
в звене детектор — командный нейрон изменяет настройку
на сигнал, не меняя структуру реакции. Пластичность
выходов командного мотонейрона и связей между первичным
и вторичными командными нейронами изменяет реакцию,
не затрагивая ее селективности в отношении вызывающего сигнала.
Управление различными реакциями происходит при
помощи поля командных нейронов, каждый из которых
запускает определенную реакцию. Нейроны связаны
между собой сильным латеральным торможением при
высоком пороге генерации сигнала. Это позволяет в каждый
данный момент обеспечивать единственность реакции.
Сложную реакцию можно «набирать» на поле командных.
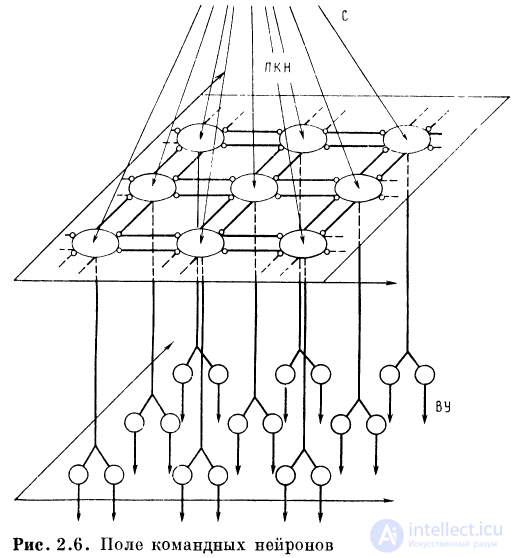
Нейронов, перемещая максимум возбуждения с нейрона
на нейрон. Управление командными нейронами можно
осуществить на основе запомненной информации —
некоторого «мнемического вектора», все компоненты которого
равны нулю, кроме одного. Последовательность
управляющих «мнемических» векторов позволяет генерировать
сложную последовательность движений на основе
механизмов памяти (рис. 2.6).
Коэффициенты связей детекторов с командными нейронами
и состояние командных нейронов определяют реакцию на
стимул. Состояние нейрона шжпо характеризовать
порогом, уровнем стационарного возоуждения и состоянием
внутреннего генератора. Состояние внутреннего
генератора пейсмекерных колебаний определяет длительную
реакцию командного нейрона на краткосрочное внешнее
воздействие. При одном и том же внешнем воздействии будет
включен командный нейрон, внутренний генератор
которого готов к работе. Изменяя состояние внутренних
генераторов командных нейронов, можно перераспределять
приоритеты реакций. Управление состоянием командных
нейронов достигается работой особого класса
модулирующих нейронов. Модулирующие нейроны, не вызывая
внешних реакций, изменяют состояние командных
нейронов, управляя готовностью внутренних генераторов.
Модулирующие нейроны можно разделить на
генерализованные и локальные. Генерализованные модулирующие
нейроны поднимают уровень готовности всех командных
нейронов. Тем самым все реакции потенциально могут
быть вызваны более слабыми стимулами. Локальные
модулирующие нейроны повышают реактивность одних и
понижают реактивность других командных нейронов,
осуществляя перераспределение приоритетов запускаемых
этими командными нейронами реакций. Модулирующий
вектор, генерируемый локальным модулирующим
нейроном, равен произведению скалярной величины его
возбуждения на вектор связей модулирующего нейрона
с командными нейронами. В результате действия
модулирующего вектора уровень реактивности одних командных
нейронов повышается, других снижается, третьих
остается без изменений.
Пластичные связи детекторов с командным нейроном могут
изменяться под влиянием проходящих через эти связи
сигналов. Изменение связей, зависящее только от
подаваемого сигнала, будем называть стимул-зависимым
обучением. Внешний сигнал генерирует на множестве
вторичных детекторов вектор возбуждения. Коэффициенты
исходных связей детекторов с командными нейронами
могут быть большими, и тогда стимул-зависимое обучение
приводит к уменьшению части коэффициентов связей,
что приводит к селективному в отношении стимула
привыканию. Если исходные связи слабые (коэффициенты
маленькие), то в результате воздействия стимула часть
связей возрастает (коэффициенты увеличиваются), что
приводит к эффекту фасилитации.
Рассмотрим эффект привыкания. Исходный вектор
связи детекторов с командными нейронами при действии
стимула умножается на оператор стимул-зависимого
обучения. Этот оператор представляется диагональной
матрицей, элементы которой представляют собой
разности единичного возбуждения и возбуждения данного
компонента вектора возбуждения, умноженного на
коэффициент обучения. Оператор стимул-зависимого обучения
по форме сходен с оператором адаптации, но относится
к вектору связи, тогда как оператор адаптации относится
к вектору возбуждения. В результате действия оператора
стимул зависимого обучения коэффициенты связей
уменьшаются тем больше, чем больше было проходящее через
эти связи возбуждение. При каждом последующем
воздействии стимула происходит умножение вектора связей
на оператор обучения. При разных сигналах операторы
обучения определяются каждый раз вектором
возбуждения вторичных детекторов. В случае слабых связей
оператор стимул-зависимого обучения состоит из
произведений коэффициента обучения на соответствующий
компонент вектора возбуждения. В результате многократного
воздействия стимула коэффициенты связей меняются
в зависимости от значений компонентов вектора
возбуждения. В случае привыкания вектор связей становится
ортогональным вектору возбуждения. Ответ командного
нейрона, равный скалярному произведению вектора
возбуждения на вектор связи, становится равным нулю. Если
сигнал изменяется, то новый вектор возбуждения не ортогонален вектору связи, и ответ командного нейрона
возрастает, демонстрируя селективность этой формы стимул-зависимого обучения.
При фасилитации вектор связи становится коллине-
арным вектору возбуждения. Реакция командного нейрона
на повторяющийся стимул достигает максимума. Если
подается новый сигнал, то вектор возбуждения перестает
быть коллинеарным сформулированному вектору связи и
реакция командного нейрона уменьшается, обнаруживая
селективность фасилитации в отношении стимула.
Особенностью стимул-зависимого привыкания является то,
что оно не связано с эффектами, следующими за реакциями.
Все изменения определяются входным сигналом.
Если опыт прерывается, то сформировавшиеся связи
возвращаются к исходному состоянию, что описывается
умножением вектора связи на оператор рассогласования,
элементы которого характеризуются коэффициентом
восстановления. Под действием оператора восстановления
привыкание и фасилитация постепенно исчезают.
Эффект-зависимое обучение характеризуется тем, что
связи между детекторами и командным нейроном
изменяются под влиянием последующего за реакцией эффекта.
В простейшем случае на командном нейроне выделяется
устойчивый вход, под влиянием которого изменяются
коэффициенты связей только тех детекторов, которые были
задействованы непосредственно перед приходом сигнала
по устойчивому входу. Эффект-зависимое обучение — это
модель классического условного рефлекса, оно имеет
место в каждом командном нейроне независимо от
характера распределения возбуждений по входам.
Следовательно, эффект-зависимое обучение распределено по
всему множеству командных нейронов, получающих из
внешней среды оценку реакции и сигнал. Изменение вектора
связей данного командного нейрона определяется тем,
какой вектор возбуждения получил подкрепление по
устойчивому входу.
Таким образом, формально эффект-зависимое обучение
можно свести к стимул-зависимому, умножая на оператор
обучения исходный вектор связей только тогда, когда за
этим вектором возбуждения следует возбуждение,
поступающее по устойчивому подкрепляющему входу. Следует различать два типа подкрепления: положительное и
отрицательное. При положительном подкреплении оператор
обучения увеличивает коэффициенты связей, при
отрицательном — уменьшает: это аналогично ситуации,
которая возникает при привыкании и фасилитации. Только
теперь при разном подкреплении используются разные
операторы обучения. Стимул, подкрепляемый
положительно, увеличивает коэффициенты связей, стимул,
подкрепляемый отрицательно, уменьшает коэффициенты
связей. В результате положительно подкрепляемый
сигнал селективно вызывает реакцию командного нейрона,
а отрицательно подкрепляемый уменьшает коэффициенты.'
В интервалах между подкреплениями исходные
значения связей восстанавливаются, что моделируется
умножением вектора связей на операторы восстановления.
Для восстановления усиленных и ослабленных связей
используются разные операторы восстановления. Кроме
стимул-зависимого и эффект-зависимого обучения,
управление связями между детекторами и командными
нейронами может быть осуществлено посредством
трансформирующего вектора, который подключает к командному
нейрону определенные детекторы. Таким образом, к
командному нейрону можно подключать разные входы.
Генерирующие векторы нейроны не вызывают реакций,
они только «переучивают» командный нейрон. Это
переучивание, однако, не требует повторений, а
осуществляется сразу без проб и ошибок. Трансформирующий вектор
изменяет коэффициенты связей на время, пока не
поступит другой трансформирующий вектор.
Обобщенное представление процесса, обучения на набор
стимулов дается матричными уравнениями. Матрица-
строка определяет вектор возбуждения, генерируемый
вторичными детекторами. В реальных условиях каждый
отдельный стимул генерирует отдельный вектор
возбуждения. Каждый командный нейрон характеризуется
вектором связей, а весь набор детекторов характеризуется
матрицей связей.
Матрица связей изменяется под влиянием поданных
сигналов путем умножения на обучающий
оператор-матрицу, элементы которой составлены из разностей или
сумм единицы и произведения коэффициента обучения на компонент вектора возбуждения. При уменьшении
связей используется разность, при увеличении — сумма.
В результате формируется новая матрица связей. При
действии следующего набора стимулов полученная ранее
матрица связей вновь умножается на оператор обучения
в результате чего возникает следующая новая матрица
связей. В отличие от стимул-зависимого обучения при
эффект-зависимом матрица связей меняется, если за
сигналом следует подкрепление. При этом направление
изменения связей зависит от знака подкрепления. Таким
образом, в ответ на одни сигналы связи возрастают, а на
другие — убывают. Восстановление связей
моделируется умножением на оператор восстановления, который
используется при отсутствии сигналов и подкрепления.
Формально принятие решения можно моделировать
возбуждением одного командного нейрона из их набора.
Выбирается нейрон, на котором достигается порог
возбуждения и одновременно — максимум реакции.
Достижение порога возбуждения — необходимое, но
недостаточное условие. Из множества достигших порога
возбуждения командных нейронов выбирается максимально
возбужденный. Выделение такого нейрона
осуществляется латеральным торможением. Степень возбуждения
каждого нейрона определяется стимул-зависимым и
эффект-зависимым обучением, влиянием
трансформирующего и вкладом поступающего от модулирующего
нейрона модулирующего вектора. В период работы
командного нейрона остальные командные нейроны находятся
в заторможенном состоянии за счет латерального
торможения.
После того как внешний сигнал представлен набором детекторов, селективно выделяющих значения определенного параметра этого сигнала, становится возможным позиционное кодирование (номером возбужденного канала) скорости изменения этого параметра. Первый этап — формирование градуальных первичных детекторов. Один первичный детектор отвечает возрастанием реакции с ростом скорости изменения, другой реагирует ослаблением реакции при увеличении скорости изменения. Из градуальных нейронов скорости изменения данного
параметра формируются вторичные детекторы, каждый из которых селективно настроен на определенное значение скорости изменения данного параметра. Организация сети первичных детекторов скорости изменения основана на использовании параллельного торможения, подаваемого с фиксированной задержкой, и определенного интервала суммации. Градуальный нейрон с малой задержкой и большим интервалом суммации имеет возрастающую
с увеличением скорости реакцию. Градуальный нейрон с большой задержкой торможения, но малым интервалом
суммации отвечает ослаблением реакции при увеличении скорости изменения параметра. Таким образом, вторичные
детекторы скорости изменения селективно настроены на разные скорости. Аналогичным образом в следующих двух слоях можно получить селективные детекторы, настроенные на разные ускорения изменения
детектируемого параметра. Детекторы направления изменения сигнала образованы за счет анизотропного латерального
торможения на входе. Благодаря такому торможению ответ элементов имеет место только при определенном
направлении изменения сигнала. В случае, когда детекторы образуют двумерное
продолжение следует...
Часть 1 2 ОБЩАЯ ТЕОРИЯ ОРГАНИЗАЦИИ СИСТЕМ ИЗ НЕЙРОПОДОБНЫХ ЭЛЕМЕНТОВ
Часть 2 2.1.16. Описание анализатора матричными уравнениями - 2 ОБЩАЯ ТЕОРИЯ ОРГАНИЗАЦИИ
Часть 3 2.1.21. возрастание различительной чувствительности под влиянием адаптации первичных детекторов -
Часть 4 - 2 ОБЩАЯ ТЕОРИЯ ОРГАНИЗАЦИИ СИСТЕМ ИЗ НЕЙРОПОДОБНЫХ ЭЛЕМЕНТОВ
Комментарии
Оставить комментарий
Нейробионика
Термины: Нейробионика