Лекция
Привет, мой друг, тебе интересно узнать все про эволюция психики элементарная сенсорная психика, тогда с вдохновением прочти до конца. Для того чтобы лучше понимать что такое эволюция психики элементарная сенсорная психика , настоятельно рекомендую прочитать все из категории Сравнительная психология и Зоопсихология.
Вводные замечания
Эволюция психики составляет часть общего процесса эволюции животного мира и совершалась по закономерностям этого процесса. Повышение общего уровня жизнедеятельности организмов, усложнение их взаимоотношений с окружающим миром приводило в ходе эволюции к необходимости все более интенсивного контактирования со все большим числом предметных компонентов среды, к совершенствованию передвижения среди этих компонентов и ко все более активному обращению с ними. Только такое прогрессивное изменение поведения могло обеспечить возрастающее потребление необходимых для жизни компонентов среды, равно как все более успешное избегание неблагоприятных или вредных воздействий и опасностей. Однако для всего этого требовалось существенное совершенствование ориентации во времени и пространстве, что и достигалось прогрессом психического отражения.
Таким образом, движение (первично локомоция, а впоследствии и манипулирование) являлось решающим фактором эволюции психики. С другой стороны, без прогрессивного развития психики не могла бы совершенствоваться двигательная активность животных, не могли бы производиться биологически адекватные двигательные реакции и, следовательно, не могло бы быть эволюционного развития.
Конечно, психическое отражение не оставалось неизменным в ходе эволюции, а само претерпевало глубокие качественные преобразования, к рассмотрению которых мы сейчас и перейдем. Существенным является то, что первично; еще примитивное, психическое отражение обеспечивало только уход от неблагоприятных условий (как это характерно и для современных простейших); поиск непосредственно не воспринимаемых благоприятных условий появился значительно позже. Как мы уже видели, такой поиск является постоянным компонентом развитого инстинктивного поведения.
Равным образом только на высоких уровнях эволюционного развития, когда уже существует предметное восприятие и сенсорные действия животных обеспечивают возникновение образов, психическое отражение становится способным полноценно ориентировать и регулировать поведение животного с учетом предметности компонентов внешней среды. Такое отражение имеет, в частности, первостепенное значение для выделения преград (в узком и широком смысле слова), что, как мы видели, является необходимым условием для появления самых лабильных форм индивидуального поведенческого приспособления к меняющимся условиям среды — навыков и, у наиболее высокоразвитых животных, интеллекта.
Признаки наиболее глубоких качественных изменений, которые претерпела психика в процессе эволюции животного мира, Леонтьев положил в основу выделенных им стадий психического развития. Четкая, наиболее существенная грань проходит между элементарной сенсорной и перцептивной психикой, знаменуя собой основную веху грандиозного процесса эволюции психики. Поэтому в дальнейшем изложении мы будем основываться на этом делении.
Элементарную сенсорную психику Леонтьев определяет как стадию, на которой деятельность животных «отвечает тому или иному отдельному воздействующему свойству (или совокупности отдельных свойств) в силу существенной связи данного свойства с теми воздействиями, от которых зависит осуществление, основных биологических функций животных. Соответственно отражение действительности, связанное с таким строением деятельности, имеет форму чувствительности к отдельным воздействующим свойствам (или совокупности свойств), форму элементарного ощущения».[48] Стадия же перцептивной психики, по Леонтьеву, «характеризуется способностью отражения внешней объективной действительности уже не в форме отдельных элементарных ощущений, вызываемых отдельными свойствами или их совокупностью, но в форме отражения вещей».[49] Деятельность животного определяется на этой стадии тем, что выделяется содержание деятельности, направленное не на предмет воздействия, а на те условия, в которых этот предмет объективно дан в среде. «Это содержание уже не связывается с тем, что побуждает деятельность в целом, но отвечает специальным воздействиям, которые его вызывают».[50]
Однако для наших целей этого подразделения недостаточно, и поэтому следует как в пределах элементарной сенсорной, так и в пределах перцептивной психики выделить существенно различающиеся уровни психического развития: низший и высший, допуская при этом существование и промежуточных уровней (рис. 25). Важно отметить, что крупные систематические таксоны животных не всегда и не вполне укладываются в эти рамки. Это неизбежно, так как в пределах крупных таксонов (в данном случае подтипов или типов) всегда имеются животные, стоящие на смежных уровнях психического развития. Объясняется это тем, что качества высшего психического уровня всегда зарождаются на предшествующем уровне.
Кроме того, расхождения между психологической и зоологической классификациями обусловлены тем, что морфологические признаки, на которых построена систематика животных, отнюдь не всегда определяют особенности и степень развития психической деятельности последних. Поведение животных представляет собой совокупность функций эффекторных органов животных. А в процессе эволюции именно функция первично определяет форму, строение организма, его систем и органов.

И лишь вторично строение эффекторов, их двигательные возможности определяют характер поведения животного, ограничивают сферу его внешней активности.
Этот диалектический процесс, однако, осложняется еще возможностями многопланового решения задач (по меньшей мере у высших животных) и компенсаторными процессами в области поведения. Это означает, что если животное в данных условиях лишено возможности решить биологически важную задачу одним путем, оно, как правило, имеет в своем распоряжении еще другие, резервные возможности. Так, одни эффекторы могут замениться другими, т. е. разные морфологические структуры могут служить для выполнения биологически однозначных действий. С другой стороны, одни и те же органы могут выполнять разные функции, т. е. осуществляется принцип мультифункциональности. Особенно гибки морфофункциональные отношения в координационных системах, прежде всего в центральной нервной системе высших животных.
Итак, с одной стороны, образ жизни определяет развитие приспособлений в эффекторной сфере, а с другой стороны, функционирование эффекторных систем, т. е. поведение, обеспечивает удовлетворение жизненных потребностей, обмена веществ в ходе взаимодействия организма с внешней средой. Изменения условий жизни порождают необходимость изменения прежних эффекторных функций (или даже появления новых функций), т. е. изменения поведения, а это затем приводит к соответствующим морфологическим изменениям в эффекторной и сенсорной сферах и в центральной нервной системе. Но не сразу и даже не всегда функциональные изменения влекут за собой морфологические. Более того, у высших животных зачастую вполне достаточными, а иногда даже наиболее результативными являются чисто функциональные изменения без морфологических перестроек, т. е. адаптивные изменения только поведения (А. Н. Северцов). Поэтому поведение в сочетании с мультифункциональностыо эффекторных органов обеспечивает животным наиболее гибкую адаптацию те новым условиям жизни.
Указанные функциональные и морфологические преобразования определяют качество и содержание психического отражения в процессе эволюции. О них, о диалектических взаимоотношениях между образом жизни (биологией) и поведением, необходимо всегда помнить, когда мы говорим о примате двигательной активности, деятельности животных, в развитии психического отражения.
Уместно еще раз напомнить и о том, что, вопреки еще бытующему представлению, врожденное и приобретаемое поведение не являются последовательными ступенями на эволюционной лестнице, а развиваются и усложняются совместно, как два компонента одного единого процесса. Следовательно, не было и нет такого положения, чтобы у низших животных имелись лишь одни инстинкты (или даже рефлексы), на смену которым у высших приходят навыки, а инстинкты все больше сходят на нет. На самом деле, как уже было показано, прогрессивному развитию именно инстинктивного, генетически фиксированного поведения соответствует прогресс в области индивидуально-изменчивого поведения. Инстинктивное поведение достигает наибольшей сложности как раз у высших животных, и этот прогресс влечет за собой развитие и усложнение у этих животных форм научения.
Мы здесь можем, разумеется, дать только самый общий обзор эволюции психики, и то лишь по некоторым его направлениям и на немногих примерах, совершенно не вскрывая всего многообразия путей развития психики в мире животных.
Глава 1
ЭЛЕМЕНТАРНАЯ СЕНСОРНАЯ ПСИХИКА
Низший уровень психического развития
На низшем уровне психического развития находится довольно большая группа животных; среди них встречаются и такие животные, которые стоят еще на грани животного и растительного мира (жгутиковые), а с другой стороны, и сравнительно сложно устроенные одноклеточные и многоклеточные животные. К наиболее типичным представителям рассматриваемой здесь группы животных относятся простейшие, на примере которых здесь будет дана характеристика низшего уровня элементарной сенсорной психики. Правда, некоторые высокоорганизованные простейшие (из числа инфузорий) поднялись уже на более высокий уровень элементарной сенсорной психики. В этой связи следует напомнить, что филогенез простейших шел в известной мере параллельно развитию низших многоклеточных животных, а это, в частности, нашло свое отражение в формировании у простейших аналогов органов таких животных. Эти аналоги получили название органелл.
Движения простейших
Движения простейших отличаются большим разнообразием, причем в этом типе простейших животных встречаются способы локомоции, которые совершенно отсутствуют у многоклеточных животных. Это своеобразный способ передвижения амеб при помощи «переливания» плазмы из одного участка тела в другой. Другие представители простейших, грегарины, передвигаются своеобразным «реактивным» способом — путем выделения из заднего конца тела слизи, «толкающей» животное вперед. Существуют и простейшие, пассивно парящие в воде.
Однако большинство простейших передвигаются активно с помощью особых структур, производящих ритмичные движения, — жгутиков или ресничек. Эти эффекторы представляют собой плазматические выросты, совершающие колебательные, вращательные или волнообразные движения. Жгутиками, длинными волосовидными выростами обладают уже упомянутые примитивные простейшие, получившие свое название благодаря этому образованию. С помощью жгутиков тело животного (например, эвглены) приводится в спиралевидное поступательное движение. Некоторые морские жгутиковые, по данным норвежского ученого И. Трондсена, вращаются при движении вокруг оси со скоростью до 10 оборотов в секунду, а скорость поступательного движения может достигнуть 370 микрон в секунду. Другие морские жгутиковые (из числа динофлягеллят) развивают скорость от 14 до 120 микрон в секунду и больше. Более сложным эффекторным аппаратом являются реснички, покрывающие в большом числе тело инфузории. Как правило, реснитчатый покров располагается неравномерно, реснички достигают на разных участках тела различной длины, образуют кольцевидные уплотнения (мембранеллы) и т. п.
Примером такой сложной дифференциации могут служить инфузории из рода стилонихия. Своеобразные органеллы этих животных позволяют им не только плавать, но и «бегать» по твердому субстрату, причем как вперед, так и назад. Установлено, что координация этих способов и направлений локомоции, как и их «переключение», осуществляется специальными механизмами, локализованными в трех центрах и двух осях градиентов возбуждения в цитоплазме.
Жгутики и реснички приводятся в движение сокращениями миофибрилл, которые образуют волоконца, мионемы, соответствующие мышцам многоклеточных животных. У большинства простейших они являются основным двигательным аппаратом, причем имеются они даже у наиболее примитивных представителей типа жгутиковых. Мионемы располагаются в строгом порядке, чаще всего в виде колец, продольных нитей или лент, а у высших представителей и в виде специализированных систем. Так, у инфузории Caloscolex имеются специальные системы мионем околоротовых мембранелл, глотки, задней кишки, ряд ретракторов отдельных участков тела и т. д.
Интересно отметить, что, как правило, мионемы имеют гомогенную структуру, что отвечает гладкой мускулатуре многоклеточных животных, однако иногда встречаются и поперечно исчерченные мионемы, сопоставимые с поперечно-полосатой мускулатурой высших животных. Все сократительные волоконца служат для выполнения быстрых движений отдельных эффекторов (у простейших — игловидных выростов, щупальцевидных образований и т. п.). Сложные системы мионем позволяют простейшим производить не только простые сократительные движения тела, но и достаточно разнообразные специализированные локомоторные и нелокомоторные движения.
У тех простейших, у которых нет мионем (у амеб, корненожек, споровиков, за одним исключением, и некоторых других простейших), сократительные движения совершаются непосредственно в цитоплазме. Так, при передвижении амебы в наружном слое цитоплазмы, в эктоплазме, происходят подлинные сократительные процессы. Удалось даже установить, что эти явления имеют место всякий раз в «задней» (по отношению к направлению передвижения) части тела амебы.
Таким образом, еще даже до появления специальных эффекторов перемещение животного в пространстве совершается путем сокращений. Именно сократительная функция, носителем которой являются у простейших мионемы, а у многоклеточных мышцы, обеспечивала все разнообразие и всю сложность двигательной активности животных на всех этапах филогенеза.
Кинезы
Локомоция простейших осуществляется в виде кинезов — элементарных инстинктивных движений. Типичным примером кинеза является ортокинез — поступательные движение с переменной скоростью. Если, например, на определенном участке существует температурный градиент (перепад температур), то движения туфельки будут тем более быстрыми, чем дальше животное будет находиться от места с оптимальной температурой. Следовательно, здесь интенсивность поведенческого (локомоторного) акта непосредственно определяется пространственной структурой внешнего раздражителя.
В отличие от ортокинеза при клинокинезе имеет место изменение направления передвижения. Это изменение не является целеустремленным, а носит характер проб и ошибок, в результате которых животное в конце концов попадает в зону с наиболее благоприятными параметрами раздражителей. Частота и интенсивность этих изменений (следовательно, и угол поворота) зависят от интенсивности воздействующего на животное (отрицательного) раздражителя (или раздражителей). С ослаблением силы действия этого раздражителя уменьшается и интенсивность клинокинеза. Таким образом, животное и здесь реагирует на градиент раздражителя, но не увеличением или уменьшением скорости передвижения, как при ортокинезе, а поворотами оси тела, т. е. изменением вектора двигательной активности.
Как мы видим, осуществление наиболее примитивных инстинктивных движений — кинезов — определяется непосредственным воздействием градиентов интенсивности биологически значимых внешних факторов. Роль внутренних процессов, происходящих в цитоплазме, заключается в том, что они дают поведенческому акту «первый толчок», как и у многоклеточных животных.
Ориентация
Уже на примерах кинезов мы видели, что градиенты внешних раздражителей выступают у простейших одновременно как пусковые и направляющие стимулы. Особенно наглядно это проявляется при клинокинезах. Однако изменения положения животного в пространстве еще не являются здесь подлинно ориентирующими, поскольку они носят ненаправленный характер. Для достижения полного биологического эффекта клинокинетические, как и ортокинетические, движения нуждаются в дополнительной коррекции, позволяющей животному более адекватно ориентироваться в окружающей его среде по источникам раздражения, а не только менять характер движения при неблагоприятных условиях.
Ориентирующими элементами являются у представителей рассматриваемого типа и у других низших беспозвоночных, стоящих на данном уровне психического развития, простейшие таксисы. В ортокинезах ориентирующий компонент — ортотаксис — проявляется в изменении скорости передвижения без изменения его направления в градиенте внешнего раздражителя. В клинокинезах этот компонент называется клинотаксисом и проявляется в изменении направления движения на определенный угол.
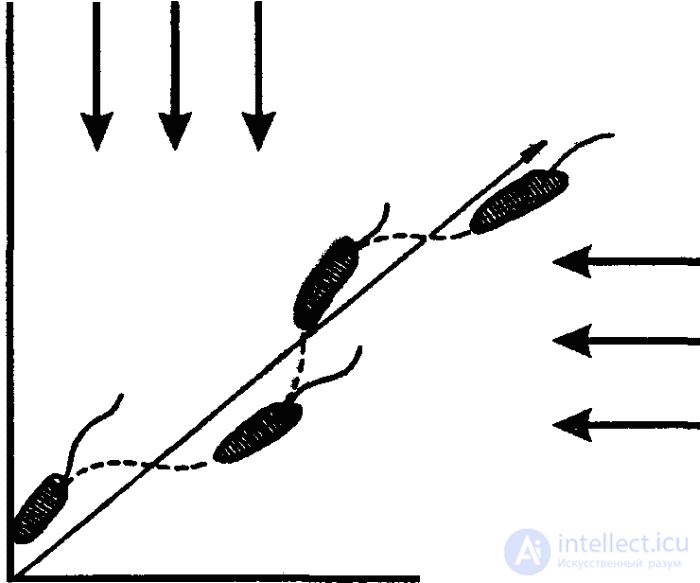
Как мы уже знаем, под таксисами понимают генетически фиксированные механизмы пространственной ориентации двигательной активности животных в сторону благоприятных (положительные таксисы) или в сторону от неблагоприятных (отрицательные таксисы) условий среды. Так, например, отрицательные термотаксисы выражаются у простейших, как правило, в том, что они уплывают из зон с относительно высокой температурой воды, реже — из зон с низкой температурой. В результате животное оказывается в определенной зоне термического оптимума (зоне предпочитаемой температуры). В случае ортокинеза в температурном градиенте отрицательный ортотермотаксис обеспечивает прямолинейное удаление от неблагоприятных термических условий. Если же имеет место клинокинетическая реакция (рис. 26), то клинотаксис обеспечивает четкое изменение направления передвижения, ориентируя тем самым случайные клинокинетические движения в градиенте раздражителя (в нашем примере — в термическом градиенте).
Зачастую клинотаксисы проявляются в ритмичных маятникообразных движениях (на месте или при передвижении) или в спиралевидной траектории плывущего животного. И здесь имеет место регулярный поворот оси тела животного (у многоклеточных животных это может быть и только часть тела, например голова) на определенный угол.
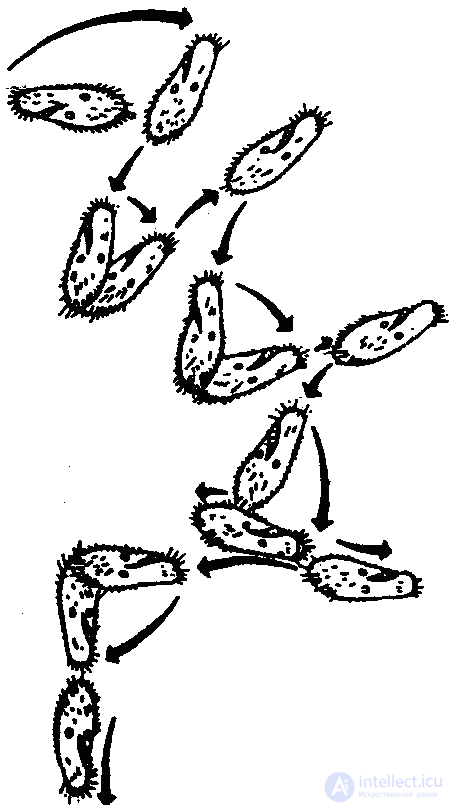
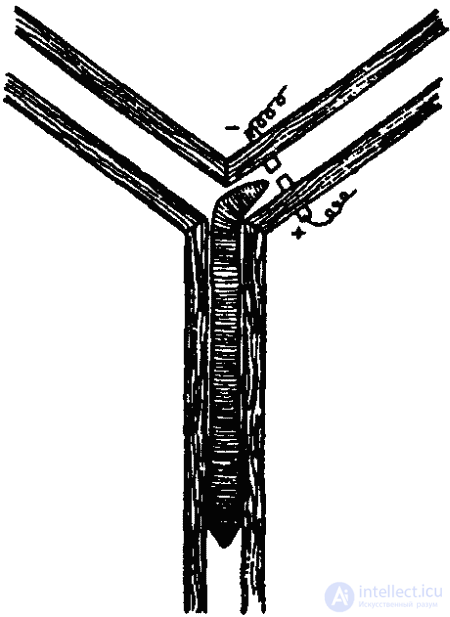
Клинотаксисы обнаруживаются и при встрече с твердыми преградами. Вот пример соответствующего поведения инфузории (туфельки). Наткнувшись на твердую преграду (или попав в зону с другими неблагоприятными параметрами среды), туфелька останавливается, изменяется характер биения ресничек, и животное отплывает немного назад. После этого инфузория поворачивается на определенный угол и снова плывет вперед. Это продолжается до тех пор, пока она не проплывет мимо преграды (или не минует неблагоприятную зону).

Согласно новым данным (В. Розе), инфузории, однако, не всегда ведут себя подобным образом. Клинотаксисы наблюдаются лишь при встрече с преградой под углом в 65–85° (рис. 27, 1). Если же встреча произойдет под прямым углом, инфузория переворачивается через поперечную ось тела и «отскакивает» назад (рис 27, 2). Если же животное встречается с преградой по касательной, оно проплывает мимо, свернув в сторону (рис. 27, 3).
Клинотаксисы хотя встречаются и у более высокоорганизованных животных, относятся к примитивным видам таксисов. Тропотаксисов у простейших нет, поскольку тропотаксисы предполагают наличие симметрично расположенных органов чувств.
В приведенных примерах описывались реакции простейших (в данном случае инфузорий) на температуру и тактильный раздражитель (прикосновение). Речь шла, следовательно, о термо- и тигмотаксисах, в последнем случае — об отрицательном тигмоклинотаксисе, возникающем в ответ на сильное тактильное раздражение (соприкосновение с твердой поверхностью объекта).
Если же, наоборот, туфелька натыкается не на твердое препятствие, а на мягкий объект (например, растительные остатки, фильтровальная бумага), она реагирует иначе: при такой слабой тактильной стимуляции инфузория останавливается и прикладывается к этой поверхности так, чтобы максимальный участок тела соприкасался с поверхностью объекта (положительный тигмотаксис). Аналогичная картина наблюдается и при воздействиях других модальностей на направление движения, т. е. положительный или отрицательный характер реакции зависит от интенсивности раздражения. Как правило, простейшие реагируют на слабые раздражения положительно, на сильные — отрицательно, но в целом простейшим больше свойственно избегать неблагоприятных воздействий, нежели активно искать положительные раздражители.
Возвращаясь к тигмотаксисам, важно отметить, что у инфузорий обнаружены специальные рецепторы тактильной чувствительности — осязательные «волоски», которые особенно выделяются на переднем и заднем концах тела. Эти образования служат не для поиска пищи, а только для тактильного обследования поверхностей объектов, с которыми животное сталкивается. Раздражение этих органелл и приводит в описанном примере к прекращению кинетической реакции.
Особенностью тигмотаксисной реакции является то, что она часто ослабевает, а затем и прекращается после прикасания к объекту максимальной поверхностью тела: приставшая к объекту туфелька в возрастающей мере начинает реагировать на иной раздражитель и все больше отделяется от объекта. Затем, наоборот, вновь возрастает роль тактильного раздражителя и т. д. В результате животное совершает возле объекта ритмичные колебательные движения.
Четко выражена у туфельки и ориентация в вертикальной плоскости, что находит свое выражение в тенденции плыть вверх (отрицательный геотаксис — ориентация по силе земного притяжения). Поскольку у парамеции не были обнаружены специальные органеллы гравитационной чувствительности, было высказано предположение, что содержимое пищеварительных вакуолей действует у нее наподобие статоцистов[51] высших животных. Обоснованность такого толкования подтверждается тем, что туфелька, проглотившая в опыте металлический порошок, плывет уже не вверх, а вниз, если над ней поместить магнит. В таком случае содержимое вакуоли (металлический порошок) уже давит не на нижнюю ее часть, а, наоборот, на верхнюю, чем, очевидно, и обусловливается переориентация направления движения животного на 180°.
Кроме упомянутых таксисные реакции установлены у простейших также в ответ на химические раздражения (хемотаксисы), электрический ток (гальванотаксисы) и др. На свет часть простейших реагируют слабо, у других же эта реакция выражена весьма четко. Так, фототаксисы проявляются у некоторых видов амеб и инфузорий в отрицательной форме, а инфузория Blepharisma снабжена даже светочувствительным пигментом, но, как правило, инфузории не реагируют на изменения освещения. Интересно отметить, что у слабо реагирующих на свет простейших отмечались явления суммации реакции на механические раздражения, если последние сочетались со световыми раздражениями. Такое сочетание может увеличить реакцию в восемь раз по сравнению с реакцией на одно лишь механическое раздражение.
В отличие от инфузорий у многих жгутиковых, особенно у эвглены, положительный фототаксис выражен весьма четко. Биологическое значение этого таксиса не вызывает сомнений, так как аутотрофное питание эвглены требует солнечной энергии. Эвглена плывет к источнику света по спирали, одновременно, как уже упоминалось, вращаясь вокруг собственной оси. Это имеет существенное значение, так как у эвглены, как я у некоторых других простейших, сильно и положительно реагирующих на свет, имеются хорошо развитые фоторецепторы. Это пигментные пятна, иногда снабженные даже отражающими образованиями, позволяющими животному локализовать световые лучи. Продвигаясь к источнику света описанным образом, эвглена поворачивает к нему то «слепую» (спинную) сторону, то «зрячую» (брюшную). И каждый раз, когда последняя (с незаслоненным участком «глазка») оказывается обращенной к источнику света, производится корректировка траектории движения путем поворота на определенный угол в сторону этого источника. Следовательно, движение эвглены к свету определяется положительным фотоклинотаксисом, причем в случае попадания ее под воздействие двух источников света попеременное раздражение фоторецептора то слева, то справа придает движению эвглены внешнее сходство с тропотаксисным поведением двусторонне-симметричных животных, обладающих парными глазами (рис. 28), о чем пойдет речь ниже.
«Глазки» описаны и у других жгутиковых. Например, массивный ярко-красный глазок имеет Collodictyon sparsevacuolata — своеобразный протист с 2–4 жгутиками и амебоидным обликом, способный быстро передвигаться как с помощью жгутиков, так и с помощью ложноножек. Особую сложность фоторецепция достигает у жгутикового Pauchetia (Dinoflagellata), у которого имеются уже аналоги существенных частей глаза многоклеточных животных, пигментное пятно снабжено не только светонепроницаемым экраном (аналог пигментной оболочки), но и све-топроницаемым образованием в форме сферической линзы (аналог хрусталика). Такой «глазок» позволяет не только локализовать световые лучи, но и собирать, в известной степени фокусировать их.
Пластичность поведения простейших
Как мы видим, и в моторной и в сенсорной сфере поведение достигает у ряда видов простейших известной сложности. Об этом говорит сайт https://intellect.icu . Достаточно указать на фобическую реакцию (реакцию испуга) туфельки в вышеописанном примере клинотаксиса: наткнувшись на твердое препятствие (или попав в иную неблагоприятную зону), туфелька резко останавливается и принимает «оборонительное положение», т. е. «съеживается», готовясь пустить в ход ядовитые стрекательные капсулы. Одновременно меняются движения ресничек, происходит тактильное и химическое обследование объекта и т. д. У эвглены фобическая реакция выражается в том, что она, остановившись, начинает производить передним концом тела круговые движения, после чего уплывает в другом направлении (рис. 29).
Ясно, что такая интеграция моторно-сенсорной активности возможна лишь с помощью специальных функциональных структур, аналогичных нервной системе многоклеточных животных. Однако о морфологии этих аналогов еще очень мало известно, и только относительно инфузории удалось с определенной достоверностью доказать существование специальной сетевидной системы проводящих путей, располагающейся в эктоплазме. Очевидно, проведение импульсов осуществляется у простейших и системой градиентов в самой цитоплазме.

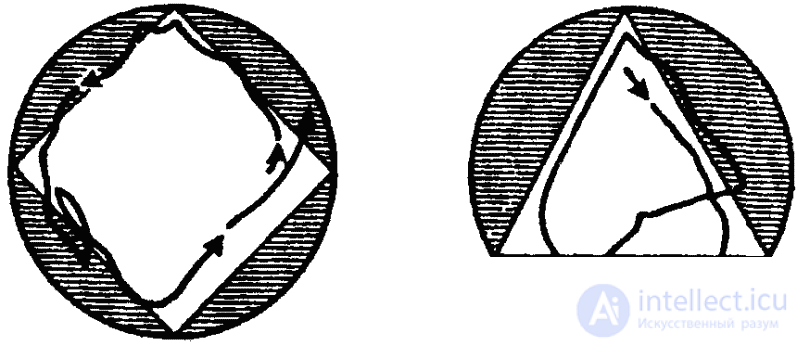
Способность к индивидуальному изменению описанных генетически фиксированных форм поведения путем научения выражена у простейших слабо. Все же способность простейших к научению, хотя бы в элементарных формах, сейчас можно считать доказанной. Если, например, держать парамеции некоторое время в треугольном или квадратном (в сечении) сосуде, то они сохраняют привычный путь передвижения вдоль стенок сосуда и после их перемещения в сосуд округлой формы (рис. 30). Аналогичным образом инфузория, плававшая длительное время (около двух часов) в сосуде, имевшем форму треугольника, придерживается этой траектории затем и в квадратном сосуде большей площади (опыты немецкого ученого Ф. Брамштедта). В опытах советской исследовательницы Н. А.Т ушмаловой инфузории были подвергнуты постоянно действующему ритмичному раздражителю — вибрации. В результате животные постепенно переставали реагировать на него обычным образом (сокращением). Исследовательница усматривает в этом пример элементарных следовых реакций, представляющих собой простую форму кратковременной памяти, формирующейся на этом уровне филогенеза за счет чисто молекулярных взаимодействий.
Подобные опыты ставились и другими экспериментаторами, применявшими различные формы воздействия. Высказывались сомнения, действительно ли в таких случаях можно говорить о каких-то формах научения, ибо сильные воздействия могли оказать хотя бы временное вредное влияние на подопытных животных. К тому же вообще в опытах с простейшими очень трудно учесть все возможные побочные факторы (особенно химические изменения среды).
Все же, очевидно, мы имеем здесь дело с элементарной формой научения — привыканием. Привыкание (к измененным внешним условиям), как мы уже знаем, играет немаловажную роль и в поведении высших животных, но имеет качественно иной характер уже хотя бы потому, что строится у них не только на ощущениях, но и на восприятиях. При элементарной же сенсорной психике животное может привыкать лишь к воздействию отдельных раздражителей (или их совокупностей), являющихся воплощением отдельных свойств или качеств компонентов окружающей среды. Именно это и имело место в приведенных примерах, когда наблюдалась модификация видотипичного поведения: врожденные реакции на определенные раздражители постепенно элиминируются, если при многократном повторении не наступит биологически значимый эффект. Так, в опытах Тушмаловой фобическая реакция у инфузорий со временем сводилась на нет при повторной встрече с «безобидным» раздражителем.
Привыкание необходимо отличать от утомления, выражающегося в явлениях «истощения» животного. В опытах Тушмаловой это выражалось в неспособности инфузории к дальнейшему реагированию, если она в течение 2–3 часов подвергалась очень сильным раздражениям. Утомление связано с перерасходом энергетических ресурсов, привыкание же представляет собой активную приспособительную реакцию, значение которой состоит в экономии этих ресурсов, в предотвращении траты энергии на бесполезные для животного движения.
Как форма научения привыкание характеризует низший уровень элементарной сенсорной психики, хотя и не теряет своего значения на всех ступенях развития психики, как это было показано при ознакомлении с онтогенезом поведения животных (см. ч. II). Однако у высших представителей простейших, возможно, уже существуют и зачатки ассоциативного научения, вообще характерного для более высоких уровней психического развития.
Об ассоциативном научении можно говорить в тех случаях, когда устанавливается временная связь между биологически значимым и «нейтральным», точнее, биологически маловалентным раздражителем. Именно такую связь Брамштедту удалось выработать у туфельки, которая, как уже отмечалось, не реагирует заметным образом на изменение освещения, но весьма чувствительна к температурным изменениям. Если в эксперименте затенить одну половину капли воды, в которой плывут парамеции, и при этом подогреть освещенную часть капли, то вскоре инфузории соберутся в холодной темной части, но останутся там (в течение 15 минут) и после того, как температура в обеих частях капли будет уравновешена. Правда, эти опыты Брамштедта подвергались серьезной критике, так как при подогреве меняется и химизм воды (меняется растворимость содержащихся в ней газов), что не может не влиять на поведение инфузории (опыты У. Грабовского).
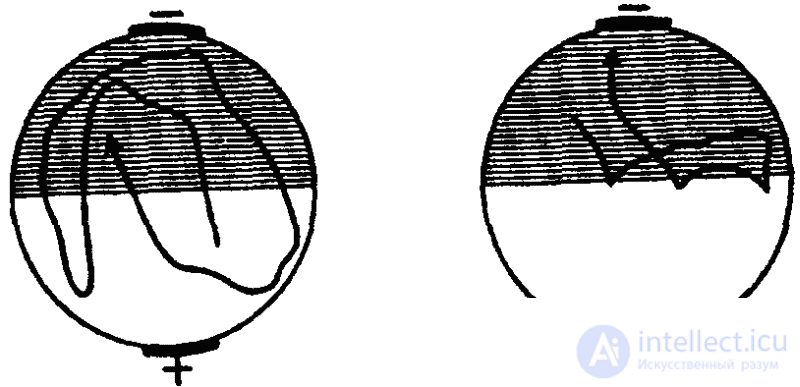
Вместе с тем аналогичный результат может быть получен и путем наказания животных при заплыве в освещенную зону ударами электрического тока (рис. 31). Эффект избегания сохранялся в этом случае в течение 20 минут (опыты Г. Зеста). Однако обратный результат, т. е. избегание темноты и заплыв в освещенную зону, Зесту получить не удалось. Это обстоятельство, однако, является результатом методической ошибки: инфузориям свойствен гальванотаксис, выражающийся в их устремлении к катоду. В упомянутых же опытах Зеста катод всегда находился на темной половине и гальванотаксис оказывался сильнее электрического раздражения, почему инфузории и направлялись только в темноту.
В более тщательно поставленных опытах польскому ученому С. Вавржинчику удалось преодолеть эти методические недостатки и с успехом научить инфузории избегать темный участок стеклянной трубки, где их раздражали электрическим током. Постепенно туфельки все чаще оставались в освещенной зоне и поворачивали на границе с затемненной зоной даже при перемещении этой границы вдоль трубки еще до получения электрического удара. Наконец, они оставались в освещенной зоне в четыре раза дольше, чем в темной, даже тогда, когда раздражения током совсем прекратились. Эта реакция сохранялась даже в течение 50 минут, что является достаточно большим сроком для простейших. Четкие положительные результаты этот исследователь получил и в других экспериментах.
Правда, Я. Дембовский, повторивший эти опыты с некоторыми изменениями, пришел к выводу, что, хотя и не исключена возможность выработки у простейших обусловленных реакций, эти опыты с туфельками все-таки не решают до конца сути вопроса, ибо существуют несравненно более сильные факторы, особенно химические, которые маскируют реакции на свет. Ввиду отсутствия вполне убедительных экспериментальных данных Дембовский оставил вопрос о наличии у простейших обусловленных реакций открытым.
Вместе с тем и у других (кроме туфельки) инфузорий, в частности у стентора и сувойки, удалось после 140–160 сочетаний выработать сократительные реакции в ответ на световое раздражение.
На инфузориях проводились и другие опыты с применением разных методических приемов. В частности, использовались капилляры, иногда с загнутым концом, в которые сажались парамеции, после чего измерялось время, необходимое им для высвобождения. В новейших исследованиях, проведенных таким способом, получались вполне положительные результаты: при каждом повторении опыта время выхода парамеции из трубочки заметно уменьшалось, что и признается доказательством их способности к ассоциативному научению.
Однако американские ученые Ф. Б. Эпплуайт и Ф. Т. Гарднер, недавно повторившие эти опыты, считают такой вывод необоснованным. Эти ученые засасывали последовательно в один и тот же капилляр разные, еще ни разу не побывавшие в опыте туфельки и установили, что время выхода этих животных из капилляра также постепенно уменьшается. Но этого не случалось, если капилляр до употребления тщательно промывался и кипятился в деионизированной воде. Использование обработанных таким образом капилляров для повторного проведения опыта с одной и той же туфелькой не давало практически никакого эффекта научения. Отсюда экспериментаторы вывели заключение, что уменьшение времени выхода из капилляра является у парамеции не результатом научения, а реакцией на загрязнение внутренней поверхности трубки продуктами обмена веществ, которое увеличивается от опыта к опыту и от которого животные пытаются все скорее уйти.
С другой стороны, в аналогичных исследованиях других ученых учитывались подобные возможные изменения среды, и было установлено, что время выхода инфузорий из трубки сокращается лишь в первых опытах, потом же оно остается постоянным (среда в трубке должна, однако, продолжать изменяться).
Таким образом, вопрос о наличии у инфузорий (а тем более у простейших вообще) ассоциативного научения еще нельзя считать решенным. Все же, очевидно, такое научение существует у них в зачаточном виде.
Общая характеристика психической активности
Как мы могли убедиться, на низшем уровне элементарной сенсорной психики поведение животных выступает в достаточно разнообразных формах, но все же мы имеем здесь дело лишь с примитивными проявлениями психической активности. О такой активности, о психике мы можем здесь говорить потому, что простейшие активно реагируют на изменения в окружающей их среде, причем реагируют на биологически непосредственно не значимые свойства компонентов среды как на сигналы о появлении жизненно важных условий среды. Другими словами, простейшим свойственна элементарная форма психического отражения — ощущение, т. е. чувствительность в собственном смысле слова. Но там, как мы уже знаем, где появляется способность к ощущению, начинается психика.
Уже говорилось и о том, что даже низший уровень психического отражения не является низшим уровнем отражения вообще, существующего в живой природе: растениям присуще допсихическое отражение, при котором имеют место лишь процессы раздражимости. Однако элементы такого допсихического отражения встречаются и у простейших.
Большой интерес представляют в этом отношении, как и вообще для понимания условий зарождения психического отражения, реакции простейших на температуру. Здесь непосредственно необходимая для поддержания жизни энергия еще тождественна опосредующей энергии, сигнализирующей о наличии жизненно необходимого компонента среды.
Еще в начале нашего века М. Мендельсон установил, что у инфузорий реакции на изменения температуры становятся все более дифференцированными по мере приближения к некому термическому оптимуму. Так, у Paramecium aurelia такой оптимум находится в пределах +24… +28° температуры воды. При +6… +15° инфузория реагирует на разности температуры от 0,06 до 0,08°, при +20… +24° же — на разности от 0,02 до 0,005°. Но по современным данным этот вид хорошо переносит и низкие температуры и даже замораживание до -30° и более.
Вместе с тем у простейших специфические терморецепторы, т. е. специальные образования, служащие для восприятия изменений температуры среды, не обнаружены. (Достоверно такие рецепторы неизвестны и для других беспозвоночных.) В свое время Г. Дженнингс полагал, что у инфузории (туфельки) передний конец тела особенно чувствителен в отношении температурных раздражений. Однако впоследствии О. Келер показал, что у разрезанной на две части туфельки передний и задний концы тела ведут себя в этом отношении в принципе так же, как целое животное. Отсюда напрашивается вывод, что реакция на изменения температуры является у простейших свойством всей протоплазмы. Возможно, что реакции простейших на термические раздражения еще сходны с биохимическими реакциями типа ферментативных процессов.
Очевидно, все это и соответствует тому обстоятельству, что для организма простейшего тепловая энергия еще не расчленяется на источник жизнедеятельности и сигнал о наличии такого источника. В связи с этим отсутствуют и специальные терморецепторы. Мы имеем, следовательно, здесь пример «сосуществования» у простейших допсихического и психического отражения.
Интересно, что отсутствие терморецепторов не делает реакции простейших менее выразительными, чем их реакции на другие внешние агенты. Как было показано, эти реакции проявляются в таксисах (в данном случае термотаксисах) и кинезах.
Еще явственнее «сосуществование» допсихического и психического отражения выступает у низших представителей типа. У эвглены оно обусловлено и наличием наряду с животным, растительного, аутотрофного типа питания, почему эвглена в одинаковой степени относится и к растениям и к животным.
С другой стороны, мы знаем, что качества психического отражения определяются тем, насколько развиты способности к движению, пространственно-временной ориентации и к изменению врожденного поведения. У простейших мы встречаем разнообразные формы передвижения в водной среде, но только на самом примитивном уровне инстинктивного поведения — кинезов. Ориентация поведения осуществляется только на основе ощущений и ограничена элементарными формами таксисов, позволяющими животному избегать неблагоприятные внешние условия. За отдельными исключениями, активность простейших находится в целом как бы под отрицательным знаком, ибо эти животные попадают в сферу действия положительных раздражителей, уходя от отрицательных.
Это значит, что поисковая фаза инстинктивного поведения (кинезов) в указанном отношении еще крайне недоразвита. К тому же она явно лишена сложной, многоэтапной структуры. Возможно, что во многих случаях эта фаза вообще отсутствует. Во всем этом проявляется не только исключительная примитивность инстинктивного поведения на данном уровне, но и предельная скудность содержания психического отражения. Ведь это содержание наполняется прежде всего активным поиском и оценкой раздражителей на первых этапах поисковой фазы.
Как уже отмечалось, в отдельных случаях у простейших встречаются и положительные элементы пространственной ориентации. К уже упомянутым примерам положительных таксисов можно добавить, что амеба в состоянии находить пищевой объект на расстоянии до 20–30 микрон. Зачатки активного поиска жертвы существуют, очевидно, и у хищных инфузорий. Однако во всех этих случаях положительные таксисные реакции еще не носят характера подлинного поискового поведения, поэтому эти исключения не меняют общую оценку поведения простейших, а тем более характеристику низшего уровня элементарной сенсорной психики в целом: дистантно на этом уровне распознаются преимущественно отрицательные компоненты среды; биологически «нейтральные» же признаки положительных компонентов, как правило, еще не воспринимаются на расстоянии как сигнальные, т. е. попросту еще не существуют для животного как таковые. Таким образом, психическое отражение выполняет на самом низком уровне своего развития в основном сторожевую функцию и отличается поэтому характерной «однобокостью»: сопутствующие биологически не значимые свойства компонентов среды дистантно ощущаются животными как сигналы появления таких компонентов лишь в том случае, если эти компоненты являются для животных вредными.
Что касается, наконец, пластичности поведения простейших, то и здесь простейшие обладают лишь самыми элементарными возможностями. Это вполне закономерно: элементарному инстинктивному поведению может соответствовать лишь элементарное научение. Последнее, как мы видели, представлено наиболее примитивной формой — привыканием, и лишь в отдельных случаях, может быть, встречаются зачатки ассоциативного научения.
Конечно, при всей своей примитивности поведение простейших является все же достаточно сложным и гибким, во всяком случае в тех пределах, которые необходимы для жизни в своеобразных условиях микромира. Эти условия отличаются рядом специфических особенностей, и этот мир нельзя себе представить как просто во много раз уменьшенный макромир. В частности, среда микромира является менее стабильной, чем среда макромира, что проявляется, например, в периодическом высыхании маленьких водоемов. С другой стороны, непродолжительность жизни микроорганизмов как отдельных особей (частая смена поколений) и относительное однообразие этого микромира делают излишним развитие более сложных форм накопления индивидуального опыта. В этой микросреде нет столь сложных и разнообразных условий, к которым можно приспособиться только путем научения. В таких условиях пластичность самого строения простейших, легкость образования новых морфологических структур в достаточной мере обеспечивают приспособляемость этих животных к условиям существования. Можно, очевидно, сказать, что пластичность поведения здесь еще не превзошла пластичность строения организма.
Как уже отмечалось, простейшие не являются однородной группой животных, и различия между их разными формами очень велики. Высшие представители этого типа во многих отношениях развивались в своеобразных формах неклеточного строения параллельно низшим многоклеточным беспозвоночным животным. В результате высокоразвитые простейшие проявляют подчас даже более сложное поведение, чем некоторые многоклеточные беспозвоночные, также стоящие на низшем уровне элементарной сенсорной психики. Это тоже одна из причин, почему мы описали этот уровень психического развития на примере только простейших. Здесь наглядно выступает уже отмеченная общая закономерность: психологическая классификация не вполне совпадает с зоологической, так как некоторые представители одной и той же таксономической категории могут еще находиться на более низком психическом уровне, другие — уже на более высоком. Именно последнее имеет место у высших представителей типа простейших, которых в этом отношении можно было бы рассматривать как исключения. Однако, в сущности, это не так, ибо здесь проявляется и другая закономерность эволюции психики, а именно: элементы более высокого уровня психического развития всегда зарождаются в недрах предшествующего, более низкого уровня. В данном случае, например, примитивные формы ассоциативного научения, вообще характерные для более высокого уровня элементарной сенсорной психики, встречаются в зачатке уже у некоторых видов, относящихся к типу, который в целом стоит на низшем уровне элементарной сенсорной психики, где типичной формой индивидуально-изменчивого поведения является привыкание.
Высший уровень развития элементарной сенсорной психики
Высшего уровня элементарной сенсорной психики достигло большое число многоклеточных беспозвоночных. Однако, как отмечалось, часть низших многоклеточных беспозвоночных находится в основном на том же уровне психического развития, что и многие простейшие. Это относится прежде всего к большинству кишечнополостных и к низшим червям, а тем более к губкам, которые во многом еще напоминают колониальные формы одноклеточных (жгутиковых). Неподвижный, сидячий образ жизни взрослых губок привел даже к редукции их внешней активности, поведения (при полном отсутствии нервной системы и органов чувств). Но даже у самых примитивных представителей многоклеточных животных создались принципиально новые условия поведения в результате появления качественно новых структурных категорий — тканей, органов, систем органов. Это и обусловило возникновение специальной системы координации деятельности этих многоклеточных образований и усложнившегося взаимодействия организма со средой — нервной системы.
К низшим многоклеточным беспозвоночным относятся помимо уже упомянутых еще иглокожие, высшие (кольчатые) черви, отчасти моллюски и др.
Мы рассмотрим в дальнейшем в качестве примера кольчатых червей, у которых в полной мере выражены признаки поведения, характерные для высшего уровня элементарной сенсорной психики. К кольчатым червям относятся живущие в морях многощетинковые черви (полихеты), малощетинковые черви (наиболее известный представитель — дождевой червь) и пиявки. Характерным признаком строения кольчецов является внешняя и внутренняя метамерия: тело состоит из нескольких большей частью идентичных сегментов, каждый из которых содержит «комплект» внутренних органов, в частности пару симметрично расположенных ганглиев с нервными коммисурами. В результате нервная система кольчатых червей имеет вид «нервной лестницы».
Нервная система
Как известно, нервная система впервые появляется у низших многоклеточных беспозвоночных. Возникновение нервной системы — важнейшая веха в эволюции животного мира, и в этом отношении даже примитивные многоклеточные беспозвоночные качественно отличаются от простейших. Важным моментом здесь является уже резкое ускорение проводимости возбуждения в нервной ткани: упротоплазме скорость проведения возбуждения не превышает 1–2 микрон в секунду, но даже в наиболее примитивной нервной системе, состоящей из нервных клеток, она составляет 0,5 метра в секунду!
Нервная система существует у низших многоклеточных в весьма разнообразных формах: сетчатой (например, у гидры), кольцевой (медузы), радиальной (морские звезды) и билатеральной. Билатеральная форма представлена у низших (бескишечных) плоских червей и примитивных моллюсков (хитон) еще только сетью, располагающейся вблизи поверхности тела, но выделяются более мощным развитием несколько продольных тяжей. По мере своего прогрессивного развития нервная система погружается под мышечную ткань, продольные тяжи становятся более выраженными, особенно на брюшной стороне тела. Одновременно все большее значение приобретает передний конец тела, появляется голова (процесс цефализации), а вместе с ней и головной мозг — скопление и уплотнение нервных элементов в переднем конце. Наконец, у высших червей центральная нервная система уже вполне приобретает типичное строение «нервной лестницы», при котором головной мозг располагается над пищеварительным трактом и соединен двумя симметричными коммисурами («окологлоточное кольцо») с расположенными на брюшной стороне подглоточными ганглиями и далее с парными брюшными нервными стволами. Существенными элементами являются здесь ганглии, поэтому говорят и о ганглионарной нервной системе, или о «ганглионарной лестнице». У некоторых представителей данной группы животных (например, пиявок) нервные стволы сближаются настолько, что получается «нервная цепочка».
От ганглиев отходят мощные проводящие волокна, которые и составляют нервные стволы. В гигантских волокнах нервные импульсы проводятся значительно быстрее благодаря их большому диаметру и малому числу синаптических связей (мест соприкосновения аксонов одних нервных клеток с дендритами и клеточными телами других клеток). Что же касается головных ганглиев, т. е. мозга, то они больше развиты у более подвижных животных, обладающих и наиболее развитыми рецепторными системами.
Зарождение и эволюция нервной системы обусловлены необходимостью координации разнокачественных функциональных единиц многоклеточного организма, согласования процессов, происходящих в разных частях его при взаимодействии с внешней средой, обеспечения деятельности сложно устроенного организма как единой целостной системы. Только координирующий и организующий центр, каким является центральная нервная система, может обеспечить гибкость и изменчивость реакции организма в условиях многоклеточной организации.
Огромное значение имел в этом отношении и процесс цефализапии, т. е. обособления головного конца организма и сопряженного с ним появления головного мозга. Только при наличии головного мозга возможно подлинно централизованное «кодирование» поступающих с периферии сигналов и формирование целостных «программ» врожденного поведения, не говоря уже о высокой степени координации всей внешней активности животного.
Разумеется, уровень психического развития зависит не только от строения нервной системы. Так, например, близкие к кольчатым червям коловратки также обладают, как и те, билатеральной нервной системой и мозгом, а также специализированными сенсорными и моторными нервами. Однако, мало отличаясь от инфузории размером, внешним видом и образом жизни, коловратки очень напоминают последних также поведением и не обнаруживают более высоких психических способностей, чем инфузории. Это опять показывает, что ведущим для развития психической деятельности является не общее строение, а конкретные условия жизнедеятельности животного, характер его взаимоотношений и взаимодействий с окружающей средой. Одновременно этот пример еще раз демонстрирует, с какой осторожностью надо подходить к оценке «высших» и «низших» признаков при сравнении организмов, занимающих различное филогенетическое положение, в частности при сопоставлении простейших и многоклеточных беспозвоночных.
Движения
Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная масса ведет придонный образ жизни в прибрежных зонах, где селится среди колоний других морских организмов или в расщелинах скал. Многие виды живут временно или постоянно в трубках, которые в первом случае периодически покидаются их обитателями, а затем вновь разыскиваются. Хищные виды отправляются из этих убежищ регулярно на «охоту». Трубки строятся из песчинок и других мелких частиц, которые скрепляются выделениями особых желез, чем достигается большая прочность построек. Неподвижно сидящие в трубках животные ловят свою добычу (мелкие организмы), подгоняя к себе и процеживая воду с помощью венчика щупалец, который высовывается из трубки, или же прогоняя сквозь нее поток воды (в этом случае трубка открыта на обоих концах).
В противоположность сидячим формам свободноживущие черви активно разыскивают свою пищу, передвигаясь по морскому дну: хищные виды нападают на других червей, моллюсков, ракообразных и иных сравнительно крупных животных, которых хватают челюстями и проглатывают; растительноядные отрывают челюстями куски водорослей; другие черви (их большинство) ползают и роются в придонном иле, проглатывают его вместе с органическими остатками или собирают с поверхности дна мелкие живые и мертвые организмы.
Малощетинковые черви ползают и роются в мягком грунте или придонном иле, некоторые виды способны плавать. Во влажных тропических лесах некоторые малощетинковые кольчецы вползают даже на деревья. Основная масса малощетинковых червей питается детритом, всасывая слизистый ил или прогрызаясь сквозь почву. Но существуют и виды, поедающие мелкие организмы с поверхности грунта, процеживающие воду или отгрызающие куски растений. Несколько видов ведут хищный образ жизни и захватывают мелких водных животных, резко открывая ротовое отверстие. В результате добыча всасывается с потоком воды.
Пиявки хорошо плавают, производя туловищем волнообразные движения, ползают, роют ходы в мягком грунте, некоторые передвигаются по суше. Помимо кровососущих существуют также пиявки, которые нападают на водных беспозвоночных и проглатывают их целиком. Наземные пиявки подстерегают свои жертвы на суше (млекопитающих и людей), в траве или на ветках деревьев и кустарников (во влажных тропических лесах). Эти пиявки могут довольно быстро двигаться. В передвижении наземных пиявок по субстрату большую роль играют присоски: животное вытягивает сперва туловище, затем присасывается к субстрату головной присоской, притягивает к ней задний конец туловища (с одновременным сокращением последнего), присасывается задней присоской и т. д.
Итак, двигательная активность кольчатых червей как при локомоции, так и при добывании пищи отличается большим многообразием и достаточной сложностью. Обеспечивается это сильно развитой мускулатурой, представленной прежде всего так называемым кожно-мышечным мешком. Он состоит из двух слоев: внешнего (подкожного), состоящего из кольцевых волокон, и внутреннего, состоящего из мощных продольных мышц. Последние простираются, несмотря на сегментацию, от переднего до заднего конца туловища. Ритмичные сокращения продольной и кольцевой мускулатуры кожно-мышечного мешка обеспечивают локомоцию: червь ползет, вытягивая и сокращая, расширяя и сужая отдельные части своего тела. Так, у дождевого червя вытягивается (и сужается) передняя часть тела, затем то же самое происходит последовательно со следующими сегментами. В результате по телу червя пробегают «волны» сокращений и расслаблений мускулатуры.
У кольчатых червей впервые в эволюции животного мира появляются подлинные парные конечности: кольчецы (помимо пиявок) носят на каждом сегменте по паре выростов, служащих органами передвижения (кроме головного конца, где они служат ротовыми органами) и получивших название параподий. Параподии снабжены специальными мышцами, двигающими их вперед или назад. Зачастую параподий имеют ветвистое строение. Каждая ветвь снабжена опорной щетинкой и, кроме того, венчиком из щетинок, имеющих у разных видов различную форму. От параподий отходят и щупальцевидные органы тактильной и химической чувствительности. Особенно длинными и многочисленными последние являются на головном конце, где на спинной стороне располагаются глаза (одна или две пары), а в ротовой полости или на особом (выпячиваемом) хоботке — челюсти. В захвате пищевых объектов могут участвовать и нитевидные щупальца на головном конце червя.
У ряда многошетинковых и всех малощетинковых червей параподии редуцированы (отсюда и название последних), остались лишь посегментно расположенные пучки щетинок. Так, у дождевого червя на каждом сегменте находится по четыре пары очень коротких, неразличимых невооруженным глазом щетинок, которые, однако, наподобие параподии служат для передвижения животного: являясь достаточно крепкими подвижными рычагами, они обеспечивают вместе с сокращениями кожно-мышечного мешка поступательное движение червя. С другой стороны, растопыривая свои щетинки и упираясь ими в грунт, дождевой червь настолько прочно фиксирует свое тело в земле, что практически невозможно вытащить его оттуда в неповрежденном виде. У некоторых других малощетинковых червей щетинки развиты значительно сильнее и представлены в большем количестве.
Органы чувств и сенсорные способности
Большой интерес для познания психической деятельности низших многоклеточных беспозвоночных представляет устройство и функционирование их органов чувств, представленных также весьма различными образованиями в соответствии с общим уровнем организации животного.
У наиболее низкоорганизованных представителей беспозвоночных органы чувств еще очень слабо дифференцированы как в морфологическом, так и в функциональном отношении. У этих животных трудно выделить органы осязания, химической чувствительности и т. д. Очевидно, первичные органы чувств вообще были плюромодальными, т. е. они обладали лишь общей, присущей всей живой материи чувствительностью, но в повышенной степени. Существование таких плюромодальных чувствительных клеток является весьма вероятной гипотезой, но в настоящее время такие рецепторные клетки, очевидно, уже не существуют. Специализация таких клеток по отдельным видам энергии привела к появлению унимодальных рецепторных образований, которые, как правило, реагируют лишь на один специфический вид энергии. Так появились термо-, хемо-, механо-, фото- и другие рецепторы.
Согласно этой гипотезе, все органы чувств многоклеточных животных развились из органов осязания — наименее дифференцированных рецепторов. В наиболее элементарных случаях осязательная функция присуща всем клеткам поверхности тела. Но уже у кишечнополостных появляются специальные осязательные клетки, которые, скапливаясь в определенных местах, образуют подлинные органы осязания. Это вытянутые цилиндрические или веретеновидные клетки, несущие на конце неподвижный чувствительный волосок или пучок волосков. Однако эти органы часто выполняют и обонятельную функцию. Особенностью низших многоклеточных беспозвоночных является то, что во многих (хотя и не во всех) случаях эти две рецепторные функции слиты и не поддаются морфологическому разграничению.
В этом нетрудно усмотреть остатки первичной плюромодальности.
С другой стороны, органы зрения относятся к наиболее сложным по строению и функционированию. Иногда органы чувств низшего порядка превращаются в органы чувств высшего порядка (например, у пиявок некоторые из органов осязания — так называемые «сенсиллы» — превращаются в глаза). Известный советский зоолог, специалист по сравнительной анатомии беспозвоночных B-А. Догель говорил в таких случаях о «повышении органа в ранге». Однако в процессе филогенеза, как отмечает Догель, нередко имело место и обратное явление; в других случаях какие-то рецепторы беспозвоночных исчезали, чтобы потом вновь появиться в несколько измененной форме. Это непостоянство и легкость перестройки привели к тому, что у близкородственных беспозвоночных однотипные рецепторы, в частности органы оптической чувствительности, подчас бывают совершенно различными по строению и функциям.
Если взять, к примеру, кишечнополостных, то гидра четко реагирует на свет, хотя специальных органов зрения у нее нет. Она воспринимает свет всей поверхностью тела. Положительный фототаксис гидры выражается в том, что животное производит в освещенной сфере круговые или маятникообразные колебательные движения и в конце концов занимает положение в сторону источника света или даже направляется (ползет) к нему. Свободноживущие же представители кишечнополостных — медузы — обладают уже специальными многоклеточными органами светочувствительности. В простейшем случае эти органы представлены так называемыми глазными пятнами, которые находятся среди обыкновенных эпителиальных клеток, и даже нечетко отграничены от них. Более дифференцированным рецептором является глазная ямка (рис. 32). Однако чаще у медуз встречаются уже настоящие глаза, причем наиболее сложно устроенные из них представляют собой погруженные под слоем эпителиальных клеток глазные пузыри приблизительно шарообразной формы. Эпителий над глазным пузырем утончен и представляет собой прозрачную роговицу. Дно и стенки пузыря состоят из двух типов клеток: ретинальных и пигментных, причем ретинальные клетки снабжены чувствительными палочками. В полости глазного пузыря находится стекловидное тело — студенистая масса, защищающая ретину от механических повреждений. Иногда встречаются даже хрусталик и радужка, и тогда налицо все основные компоненты глаза высших животных (нет, однако, глазодвигательных мышц и систем фокусировки).
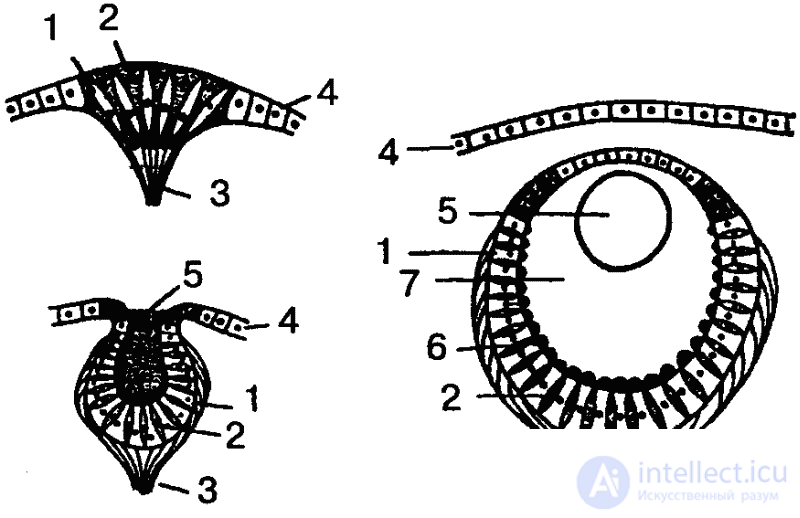
Учитывая, что глазное пятно является исходной формой вообще всех органов зрения, можно, следовательно, в ряду медуз проследить путь усложнения структуры от самого примитивного органа светочувствительности до сложного, высокодифференцированного глаза.
Очень разнообразны по своему строению и глаза червей, как и других низших многоклеточных беспозвоночных; в принципе к ним относится то же, что говорилось в отношении глаз медуз. В соответствии с многообразием движений кольчатых червей, разнообразием способов добывания пищи и других моментов жизнедеятельности находится и уровень развития сенсорной сферы этих животных. Это, правда, не означает, что у кольчецов имеются рецепторы для всех видов энергии, воздействующих на них, или даже что для всех форм чувствительности имеются специальные органы чувств. Так, например, у кольчатых червей встречаются сложно устроенные глаза, снабженные даже хрусталиками. Имеются весьма сложные глаза и у некоторых планарий и улиток. По их обладатели, насколько известно, неспособны к зрительному восприятию предметов. За исключением, может быть, некоторых улиток, у всех этих животных фотоскопические глаза, позволяющие отличать свет от тьмы и направление, откуда световые лучи падают на животное, а также перемещение светотеней в непосредственной близости от животного. Светочувствительность может при этом быть очень высокой, например, гребешок, двустворчатый моллюск с несколькими десятками глаз, закрывает створки раковины уже при уменьшении интенсивности освещения на 0,3 %. (Для сравнения можно указать, что человек воспринимает уменьшение освещения лишь не менее чем на 1 %).
Большой интерес представляют активно плавающие многощетинковые кольчатые черви из семейства Alciopidae, ведущие хищный образ жизни. У этих полихет глаз не только отличается исключительно сложным строением и величиной, но и снабжен аккомодационным устройством в виде специальных сократительных волокон, способных передвигать хрусталик и тем самым менять фокусное расстояние. Это единственный известный случай среди низших беспозвоночных: аккомодация глаза встречается только у головоногих моллюсков и позвоночных. Возможно, у этих червей в какой-то степени уже существует предметное зрение, что было бы исключением, подтверждающим общее правило. Это относится и к свободно плавающим хищным моллюскам Heteropoda, которые тоже обладают весьма сложно устроенными глазами с приспособлением, заменяющим аккомодацию.
Что же касается дождевого червя, то здесь обнаруживается чрезвычайно интересный факт: у него нет не только сложно устроенных, но и вообще никаких специальных органов светочувствительности. Вместе с тем ему свойствен четкий отрицательный фототаксис. Функцию светоощущения выполняют рассеянные в коже светочувствительные клетки. Это пример кожной светочувствительности низших многоклеточных беспозвоночных. Кожная светочувствительность наблюдается и у многих моллюсков, причем у двустворчатых это нередко единственная форма фоторецепции. Эти моллюски реагируют как на освещение, так и на затемнение чаще всего одинаковым образом — втягиванием выступающих из раковин частей тела или запиранием раковины. Многие улитки реагируют на внезапное затемнение сокращением «ноги», причем эта реакция сохраняется и после экстирпации глаз, что опять-таки указывает на наличие кожной светочувствительности.
Реакция дождевого червя на условия освещения состоит в том, что он уползает в зону большего затемнения. Если же внезапно уменьшить интенсивность освещения, то червь реагирует на это движениями бегства; в естественных условиях он вползает в почву. Такая же реакция следует за внезапным освещением. Если же осветить лишь определенный участок тела дождевого червя, то время этой реакции сокращается пропорционально размеру освещаемой площади поверхности тела — в наибольшей мере червь реагирует при освещении всей его поверхности. Следовательно, реакция определяется градиентом раздражения освещенных и неосвещенных частей тела. Аналогично реагируют на свет и другие кольчатые черви.
Кольчатые черви реагируют также на прикосновения, химические и термические раздражения, силу тяжести, электрические раздражения, течение воды, а наземные формы (дождевые черви) — на влажность среды. Однако принципиальных отличий от реакции на свет нет: все эти реакции находятся на одном уровне и характеризуются тем, что являются ответами на отдельные раздражители, на отдельные признаки, качества предметов, но не на сами предметы как таковые. Так, например, много- и малощетинковые черви проявляют четко выраженные таксисные реакции на тактильные раздражения. Преобладают отрицательные реакции, но в ряде случаев наблюдаются и положительные тигмотаксисы: прикосновения нервных сегментов животного к субстрату влекут за собой прижимание к нему всем телом, что имеет, конечно, большое значение при роющем образе жизни или жизни в трубках. Интересно, что у дождевого червя соответствующие рецепторные образования представлены лишь отдельными чувствительными клетками, разбросанными по всему телу, но более густо располагающимися на его переднем конце. У многощетинковых же червей органами осязания часто являются щупальца или щетинки.
Хорошо развита и химическая чувствительность, причем в большинстве случаев наблюдается отрицательный хемотаксис. При большой интенсивности химического воздействия черви всегда реагируют отрицательно. С другой стороны, дождевой червь, например, способен по химическим признакам выбирать разные виды листьев, что свидетельствует о специализации в сенсорной сфере. У ряда кольчецов обнаружены расположенные около ротового отверстия органы химической чувствительности в виде ямок. Особенно это относится к пларающим видам, у которых имеется пара таких ямок, выстланных мерцательным эпителием.
Таксисы
Как и на более низких уровнях эволюционного развития, пространственная ориентация совершается на высшем уровне элементарной сенсорной психики, преимущественно на основе примитивных таксисов. Но с усложнением жизнедеятельности организмов возрастают и требования к локализации биологически значимых компонентов среды по их биологически незначимым признакам. Возникает необходимость более сложного таксисного поведения, позволяющего животному достаточно четко и дифференцированно ощущать и реагировать. Именно этим и отличаются животные, находящиеся на рассматриваемом здесь уровне развития психики: благодаря симметричному расположению органов чувств у них обнаруживаются наряду с кинезами и элементарными таксисами и некоторые высшие формы таксисного поведения.
Немецкий ученый А. Кюн выделил следующие категории высших таксисов, которые, правда, вполне развиты лишь у высших животных: тропотаксисы — движение ориентируется по равнодействующей, образуемой в результате выравнивания интенсивности возбуждения в симметрично расположенных рецепторах; телотаксисы — выбор и фиксация одного источника раздражения и направление движения к этому источнику («цели»); менотаксисы — при несимметричном раздражении в симметрично расположенных рецепторах движение производится под углом к источнику раздражения. Менотаксисы играют ведущую роль в сохранении животным константного положения в пространстве.
Кроме того, у высших животных с развитой памятью встречается еще одна форма таксисов — мнемотаксисы, при которых основную роль играет индивидуальное запоминание ориентиров, что особенно важно для территориального поведения, о чем еще пойдет речь.
В поведении кольчецов тропо- и телотаксисы чаще всего проявляются совместно: если животное подвергнуть одновременному воздействию двух источников энергии (например, света) одинаковой силы, то при положительном таксисе оно будет первоначально двигаться по направлению равнодействующей, т. е. линии, ведущей к середине расстояния между источниками (тропотаксис), но затем, как правило, еще не дойдя до этой точки, оно повернет и направится к одному из источников энергии (телотаксис).
Наиболее четко тропо- и телотаксисы проявляются у активно двигающихся кольчатых червей, прежде всего у хищников. Последние не только обнаруживают присутствие жертвы по исходящим от нее физическим и химическим стимулам, но и направляются к ней, ориентируясь на основе таких таксисов. Наземные пиявки, например, способны точно локализовать местонахождение жертвы и с удивительной быстротой направляются к ней, ориентируясь по сотрясению субстрата, а затем, на более близком расстоянии, и по исходящему от жертвы (крупных млекопитающих) теплу и запаху. В результате хищник обнаруживает ее по соответствующим градиентам с помощью сочетания положительных вибрационных, термических и химических телотаксисов.
При менотаксисах животное, как говорилось, двигается под углом к линии воздействия, исходящей от источника энергии. Типичный пример — ориентация мигрирующих животных по солнечному (или другому астрономическому) «компасу», что, конечно, особенно характерно для перелетных птиц. У низших многоклеточных беспозвоночных менотаксисы обнаружены у некоторых улиток. Если этих животных поместить на освещаемый сбоку и медленно вращающийся диск, то они, двигаясь против направления вращения, придерживаются определенного угла по отношению к источнику света. В этих случаях, следовательно, можно уже говорить об ориентации животного по световому компасу, которая играет исключительную роль в жизни высших животных.
Зачатки высших форм поведения
Многощетинковые кольчатые черви относятся к наиболее развитым в психическом отношении низшим беспозвоночным. Их поведение отличается подчас большой сложностью и представляет особый интерес в том смысле, что содержит ряд элементов психической деятельности, присущей более высокоорганизованным животным. В отличие от основной массы низших беспозвоночных у полихет наблюдаются некоторые существенные усложнения видотипичного поведения, отчасти уже выходящие за рамки типичной элементарной сенсорной психики.
Так, в некоторых случаях у этих морских червей встречаются действия, которые уже можно назвать конструктивными, поскольку животные активно создают сооружения из отдельных посторонних частиц, скрепляя их в единое целое. Речь идет о постройке уже упомянутых «домиков» — трубок из отдельных частичек, которые собираются червями на морском дне и укрепляются с помощью специальных «рабочих» органов — преобразованных передних параподий.
Сам процесс сооружения трубки является сложной деятельностью, состоящей из нескольких фаз, адекватно видоизменяющихся в зависимости от таких внешних факторов, как характер грунта и течения, рельеф дна, количество и состав опускающихся на дно частиц, скорость их оседания и т. д. Строительным материалом служат разного рода мелкий обломочный материал (например, кусочки раковин), песчинки, частицы растений и т. п., которые скрепляются специальными выделениями особого «цементирующего органа». Пригодные для употребления частицы выбираются животным, причем в зависимости от его возраста; молодые особи используют лишь мелкие гранулы, более старые — более крупные. Все частицы собираются и прикрепляются друг к другу уже упомянутыми специализированными щупальцами. Полихета Aulophorus carteri, например, строит трубки из спор водных растений, которые она собирает и прикрепляет друг к другу вокруг себя подобно тому, как строится труба из кирпичей (рис. 33).

Не меньший интерес представляют впервые намечающиеся у многощетинковых червей проявления брачного поведения и агрессивности, а вместе с ними и элементов общения. Конечно, подлинные агрессивность и брачное поведение характеризуются ритуализацией, которая появляется только у головоногих моллюсков и членистоногих, т. е. на низшем уровне перцептивной психики. Все же у представителей рода Nereis удалось наблюдать борьбу между двумя червями. У Nereis pelagica такая борьба может начаться (но не обязательно), если одна особь пытается проникнуть в «домик» другой особи. Иногда вторгшаяся особь кусает владельца «домика» в задний конец тела, и тогда «хозяин» покидает трубку или же, перевернувшись, вступает в борьбу с пришельцем. В других случаях, когда животные встречаются в трубке на полпути, головой к голове, борьба вспыхивает немедленно, и черви борются, расположившись брюшными сторонами и головами друг к другу. Однако при всех обстоятельствах животные не наносят друг другу повреждения. В более ожесточенной, «непримиримой» форме борьба происходит у Nereis caudata, причем не только из-за обладания «домиком». «Драки» описаны английским исследователем поведения морских кольчатых червей С. М. Эвансом и его сотрудниками также у полихеты Harmothoe imbricata. У этого вида борьба может возникнуть при случайной встрече двух особей, особенно же агрессивность проявляется в сфере воспроизведения, при образовании пар: самец, уцепившийся за самку, становится чрезвычайно агрессивным по отношению к другим самцам (но не самкам). Никогда, однако, борьба не сопровождается у полихет подачей каких-либо сигналов или другими проявлениями ритуализации поведения.
Виноградные улитки же выполняют сложные «брачные игры», длящиеся иногда по нескольку часов, во время которых партнеры принимают по отношению друг к другу различные позы, колют друг друга известковыми спикулами («любовными стрелами») и т. д. Только после такой взаимной стимуляции начинается собственно спаривание (перенос сперматофора). У некоторых полихет, например у Platynereis dumerilii, в последние годы также были описаны брачные «танцы», однако, очевидно, еще рано делать выводы о их конкретном значении и о том, имеют ли они какое-то отношение к ритуализованному поведению. Но во всех этих случаях, несомненно, обнаруживаются некие зачатки, предваряющие значительно более сложные формы поведения высших позвоночных.
Пластичность поведения
Поведение кольчатых червей, как и других низших беспозвоночных, характеризуется малой пластичностью, консервативностью. Господствуют врожденные стереотипы («врожденные программы поведения»). Индивидуальный опыт, научение, играет в жизни этих животных еще небольшую роль. Ассоциативные связи формируются у них с трудом и лишь в ограниченных пределах. Результаты научения сохраняются недолго. У всех кольчатых червей встречается наиболее простая форма научения — привыкание, с которым мы уже встречались у простейших, у которых она является главной, если не единственной формой модификации врожденного, видотипичного поведения. У низших многоклеточных животных врожденные реакции на определенные раздражения также прекращаются после многократного повторения, если не последует адекватного подкрепления этих реакций. Так, например, дождевые черви перестают реагировать на повторное затенение, если оно остается без последствий.
Отмечается привыкание и в сфере пищевого поведения: если многощетинкового червя повторно «кормить» комочками бумаги, смоченными соком его обычной жертвы, он перестает их принимать. Если попеременно с такими комочками давать ему подлинные кусочки пищи, то он в конце концов научается различать их и будет отвергать лишь несъедобную бумагу. Подобные опыты ставились и на кишечнополостных (на полипах, находящихся, как мы уже знаем, на более низком уровне элементарной сенсорной психики). При этом полипы вели себя так же, как морские черви: уже после нескольких (до 5) опытов они отбрасывали несъедобные объекты еще до того, как подносили их к ротовому отверстию. Интересно, что нетренированные щупальца этого не делали, даже находясь рядом с тренированными.
Эти эксперименты интересны тем, что показывают способность низших беспозвоночных отличить по побочным физическим качествам съедобное от несъедобного (по вкусу предлагавшиеся объекты были одинаковыми), что подтверждает наличие истинного психического отражения уже на этом, низшем, уровне филогенетического развития. Ведь здесь производится опосредствованное действие, отличающееся тем, что свойство (или сочетание нескольких свойств), которым животное руководствуется при оценке пригодности объекта к пищевому употреблению, выступает как подлинный сигнал, а чувствительность червя или полипа играет роль посредника между организмом и компонентом среды, от которого непосредственно зависит существование животного.
Более сложное научение путем «проб и ошибок» и формирования новой индивидуальной двигательной реакции можно в элементарной форме обнаружить, начиная уже с плоских червей. Так, планарии, встречая на своем пути полоску наждачной бумаги, сперва останавливаются, но затем все же переползают через нее. Если же сочетать соприкосновение червя с наждачной бумагой с его сотрясением, то планарии перестают переползать через нее и тогда, когда не производится сотрясения. Правда, здесь, очевидно, еще нет подлинной ассоциации между двумя раздражителями (шероховатость и сотрясение); скорее всего, происходит суммация первоначально не очень сильного отрицательного раздражения (шероховатость) с дополнительным отрицательным раздражением (сотрясение) и что, вероятно, влечет за собой общее повышение возбудимости животного.
Все же можно выработать уже у планарии и более сложные реакции, когда один из раздражителей является биологически «нейтральным». В таких случаях можно говорить, очевидно, уже об элементарных процессах подлинного ассоциативного научения. Так, например, Л. Г. Воронин и Н. А. Тушмалова сумели выработать у планарии и кольчатых червей оборонительные и пищевые условные рефлексы. При этом были получены и данные, свидетельствующие об усложнении форм временных связей. Если по поводу плоских червей можно говорить лишь о примитивных неустойчивых условных рефлексах, то у многощетинковых червей обнаруживаются уже стабильные самовосстанавливающиеся после угасания условные рефлексы. Эти различия соответствуют глубоким морфологическим различиям в строении нервной системы этих групп животных и свидетельствуют о существенном прогрессе психической активности у полихет.
Пластичность поведения дождевых червей очень убедительно была показана еще в 1912 г. известным американским зоопсихологом Р. Иерксом. В проведенных им опытах черви должны были в Т-образном лабиринте выбирать определенную сторону, где находилось «гнездо» (в противоположной стороне червь получал электрический удар). Чтобы этому научить червей, потребовалось 120–180 опытов. (Улитки осваивают такую задачу после 60 опытов и запоминают верное решение в течение 30 дней.) Впоследствии аналогичные опыты с дождевыми червями ставились Л. Геком, И. С. Робинсоном, Р. И. Войтусяком (рис. 34) и другими исследователями. Удавалось также переучивать червей, поменяв «гнездо» и электроды местами. Характерно, что результат научения не меняется и после удаления передних сегментов. Более того, даже лишенный с самого начала этих сегментов, червь оказывается способным научиться правильно ориентироваться в лабиринте.
Подобные факты побудили в свое время В. А. Вагнера говорить о «сегментарной психологии» кольчатых червей и других «членистых», имея при этом в виду, что ганглии каждого сегмента в большой степени обеспечивают автономное выполнение элементарных психических функций. Обезглавленные кольчецы, писал Вагнер, «не теряют способности к спонтанным движениям, за исключением тех лишь, которые стоят в прямой зависимости и связи с органами чувств головы… Обезглавленные черви удерживают свои инстинктивные действия, даже те, в которых голова принимает прямое участие».[52]
Все это свидетельствует о том, что на уровне даже высших червей цефализация еще не достигла такого развития, чтобы оказать решающее влияние на все поведенческие акты животного. Все же общая направляющая роль головного мозга достаточно велика уже и на этом уровне. Это проявляется, например, в том, что если разрезать пополам дождевого червя, передняя половина будет закапываться в почву, производя вполне координированные движения, задняя же будет лишь беспорядочно извиваться.
Еще лучше, чем у малощетинковых червей, как уже отмечалось, вырабатываются ассоциативные связи у полихет. В опытах удавалось, например, переделать отрицательную реакцию на освещение в положительную путем сочетания освещения с подкармливанием. В итоге черви стали при освещении выползать из своих домиков и без пищевого подкрепления. Время реакции (между световым раздражением и ответным движением) уменьшилось при этом с 28,9 до 3,5 секунды (опыты М. Купленда).
Обстоятельно изучал способность к научению полихет С. М. Эванс. Применяя наказания электрическим током, этот исследователь довольно легко выработал у подопытных животных отрицательную двигательную реакцию (отказ от вползания в узкий темный коридор). В других опытах червям приходилось проползать Т-образный лабиринт. При правильном повороте они могли в течение 5 минут пребывать в затемненной камере. Особенно хорошо обучались полихеты Nereis virens, которым понадобилось менее 100 сочетаний для почти безошибочного решения задачи и ее прочного запоминания; спустя 48 часов они безошибочно воспроизводили правильный путь через лабиринт (рис. 35).
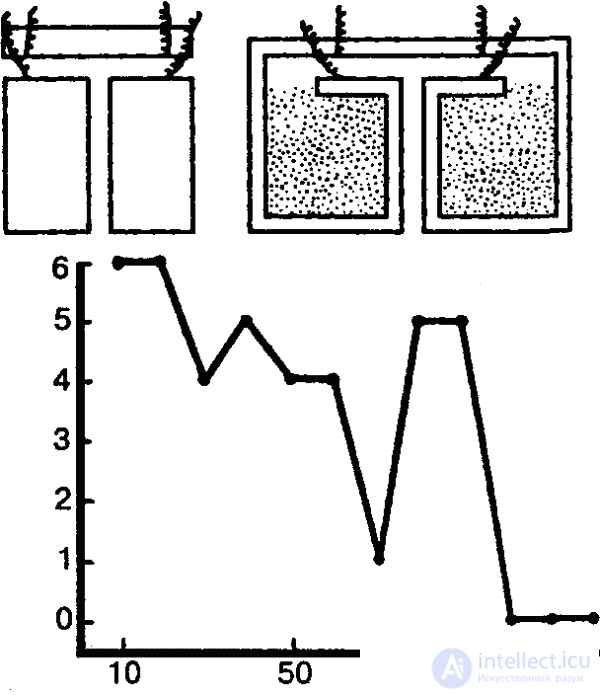
Общая характеристика высшего уровня элементарной сенсорной психики
Как уже отмечалось, наиболее низкоорганизованные формы многоклеточных беспозвоночных стоят на том же уровне психического развития, что и высшие представители простейших. Но то, что здесь не обнаруживаются существенные различия в поведении, несмотря на глубокие различия в строении, не должно нас удивлять, ибо, как уже говорилось, простейшие олицетворяют собой совершенно особую, рано отклонившуюся филогенетическую ветвь, которая до известных пределов развивалась параллельно ветви низших многоклеточных животных.
Что же касается поведения кольчатых червей, то оно вполне отвечает стадии элементарной сенсорной психики, ибо слагается из движений, ориентированных лишь по отдельным свойствам предметов и явлений (или их сочетаниям), причем это те свойства, которые, по Леонтьеву, оповещают о появлении жизненно важных условий среды, от которых зависит осуществление основных биологических функций животных. Эта ориентация осуществляется, таким образом, на основе одних лишь ощущений. Перцепция, способность к предметному восприятию, еще отсутствует. Не исключено, правда, что у некоторых наземных улиток, как и у упомянутых выше свободно плавающих хищных моллюсков и полихет, уже намечаются зачатки этой способности. Так, виноградная улитка обходит преграду еще до прикосновения к ней, ползет вдоль нее, но только если преграда не слишком велика; если же изображение преграды занимает всю сетчатку, улитка наталкивается на нее. Не реагирует она и на слишком мелкие предметы.
В поведении кольчатых червей еще преобладает избегание неблагоприятных внешних условий, как это имеет место у простейших. Но все же активный поиск положительных раздражителей занимает в поведении кольчецов уже заметное место, и это весьма характерно для высшего уровня элементарной сенсорной психики. Так же как и у простейших, в жизни кольчатых червей и других низших многоклеточных беспозвоночных большую роль играют кинезы и элементарные таксисы. Но наряду с ними здесь уже встречаются зачатки сложных форм инстинктивного поведения (особенно у некоторых многощетинковых червей, пиявок, а также улиток) и впервые появляются высшие таксисы, обеспечивающие значительно более точную и экономную ориентацию животного в пространстве, а тем самым и более полноценное использование пищевых ресурсов в окружающей среде. В результате возникли предпосылки для поднятия всей жизнедеятельности на более высокую ступень, что и имело место на стадии перцептивной психики.
Как было показано, у высших представителей рассматриваемой группы беспозвоночных впервые появляются зачатки конструктивной деятельности, агрессивного поведения, общения. Здесь опять подтверждается правило, что высшие формы поведения зарождаются уже на низших стадиях развития психической деятельности.
Давая общую оценку поведению низших многоклеточных беспозвоночных, следует отметить, что, очевидно, первично главная функция еще примитивной нервной системы состояла в координации внутренних процессов жизнедеятельности в связи со все большей специализацией клеток и новых образований — тканей, из которых строятся все органы и системы многоклеточного организма. «Внешние» же функции нервной системы определяются степенью внешней активности, которая у этих животных находится на еще невысоком уровне, зачастую не более высоком, чем у высших представителей простейших. Вместе с тем строение и функции рецепторов, как и «внешняя» деятельность нервной системы, значительно усложняются у животных, ведущих более активный образ жизни. Особенно это относится к свободноживущим, активно передвигающимся формам.
Говоря об обширной и пестрой по составу группе низших беспозвоночных, нужно, конечно, учесть, что поведение этих животных еще очень слабо изучено. Мы, например, еще почти ничего не знаем об онтогенезе поведения этих животных, о том, как формируется и развивается их поведение, да и совершенствуется ли оно вообще в процессе индивидуального развития, на что уже указывалось в части, посвященной онтогенезу поведения. Вполне возможно (если, конечно, исключить метаморфозные преобразования, личиночное поведение у низших многоклеточных и т. п.), что подобное онтогенетическое совершенствование у обсуждаемых животных не является существенным или даже вообще не происходит, ибо жесткость врожденных программ поведения, исключительная стереотипия форм реагирования являются у этих животных определяющей чертой всего их поведения.
Если я не полностью рассказал про эволюция психики элементарная сенсорная психика? Напиши в комментариях Надеюсь, что теперь ты понял что такое эволюция психики элементарная сенсорная психика и для чего все это нужно, а если не понял, или есть замечания, то не стесняйся, пиши или спрашивай в комментариях, с удовольствием отвечу. Для того чтобы глубже понять настоятельно рекомендую изучить всю информацию из категории Сравнительная психология и Зоопсихология
Комментарии
Оставить комментарий
Сравнительная психология и Зоопсихология
Термины: Сравнительная психология и Зоопсихология